Orexin
Orexin (/ɒˈrɛksɪn/), also known as hypocretin, is a neuropeptide that regulates arousal, wakefulness, and appetite.[1] The most common form of narcolepsy, in which the sufferer experiences brief losses of muscle tone (cataplexy), is caused by a lack of orexin in the brain due to destruction of the cells that produce it.[2][3]
| Prepro-orexin | |||||||||
|---|---|---|---|---|---|---|---|---|---|
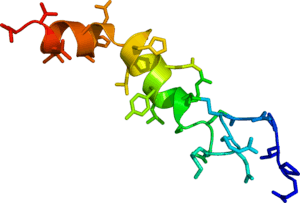 Solution phase NMR structure of orexin A based on the PDB coordinates 1R02. | |||||||||
| Identifiers | |||||||||
| Symbol | Orexin | ||||||||
| Pfam | PF02072 | ||||||||
| InterPro | IPR001704 | ||||||||
| SCOPe | 1cq0 / SUPFAM | ||||||||
| OPM superfamily | 145 | ||||||||
| OPM protein | 1wso | ||||||||
| |||||||||
| orexin (hypocretin) neuropeptide precursor | |
|---|---|
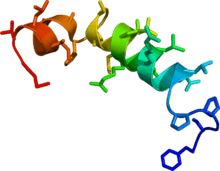 Solution phase NMR structure of orexin B based on the PDB coordinates 1CQ0. | |
| Identifiers | |
| Symbol | HCRT |
| Alt. symbols | PPOX, OX |
| NCBI gene | 3060 |
| HGNC | 4847 |
| OMIM | 602358 |
| RefSeq | NM_001524 |
| UniProt | O43612 |
| Other data | |
| Locus | Chr. 17 q21 |
There are only 10,000–20,000 orexin-producing neurons in the human brain,[2] located predominantly in the perifornical area and lateral hypothalamus.[1][4] They project widely throughout the central nervous system, regulating wakefulness, feeding, and other behaviours.[1] There are two types of orexin peptide and two types of orexin receptor.[5][4]
Orexin was discovered in 1998 almost simultaneously by two independent groups of researchers working on the rat brain.[6][7] One group named it orexin, from orexis, meaning "appetite" in Greek; the other group named it hypocretin, because it is produced in the hypothalamus and bears a weak resemblance to secretin, another peptide.[2] Officially, hypocretin (HCRT) is used to refer to the genes and transcripts, while orexin is used to refer to the encoded peptides.[8] There is a high affinity between the orexin system in the rat brain and that in the human brain.[5]
Discovery
In 1998, reports of the discovery of orexin/hypocretin were published nearly simultaneously. Luis de Lecea, Thomas Kilduff, and colleagues reported the discovery of the hypocretin system at the same time as Takeshi Sakurai from Masashi Yanagisawa's lab at the University of Texas Southwestern Medical Center at Dallas reported the discovery of the orexins to reflect the orexigenic (appetite-stimulating) activity of these peptides. In their 1998 paper describing these neuropeptides, they also reported discovery of two orexin receptors, dubbed OX1R and OX2R.[6]
The two groups also took different approaches towards their discovery. One team was interested in finding new genes that were expressed in the hypothalamus. In 1996, scientists from the Scripps Research Institute reported the discovery of several genes in the rat brain, including one they dubbed "clone 35." Their work showed that clone 35 expression was limited to the lateral hypothalamus.[9] They extracted selective DNA found in the lateral hypothalamus. They cloned this DNA and studied it using electron microscopy. Neurotransmitters found in this area were oddly similar to the gut hormone, secretin, a member of the incretin family, so they named hypocretin to stand for a hypothalamic member of the incretin family.[10] These cells were first thought to reside and work only within the lateral hypothalamus area, but immunocytochemistry tactics revealed the various projections this area truly had to other parts of the brain. A majority of these projections reached the limbic system and structures associated with it (including the amygdala, septum, and basal forebrain area).
On the other hand, Sakurai and colleagues were studying the orexin system as orphan receptors. To this end, they used transgenic cell lines that expressed individual orphan receptors and then exposed them to different potential ligands. They found that the orexin peptides activated the cells expressing the orexin receptors and went on to find orexin peptide expression specifically in the hypothalamus. Additionally, when either orexin peptide was administered to rats it stimulated feeding, giving rise to the name 'orexin'.[6]
The nomenclature of the orexin/hypocretin system now recognizes the history of its discovery. "Hypocretin" refers to the gene or genetic products and "orexin" refers to the protein, reflecting the differing approaches that resulted in its discovery. The use of both terms is also a practical necessity because "HCRT" is the standard gene symbol in databases like GenBank and "OX" is used to refer to the pharmacology of the peptide system by the International Union of Basic and Clinical Pharmacology.[8]
Isoforms
There are two types of orexin: orexin-A and -B (hypocretin-1 and -2). They are excitatory neuropeptides with approximately 50% sequence identity, produced by cleavage of a single precursor protein. Orexin-A is 33 amino acid residues long and has two intrachain disulfide bonds; orexin-B is a linear 28 amino acid residue peptide. Although these peptides are produced by a very small population of cells in the lateral and posterior hypothalamus, they send projections throughout the brain. The orexin peptides bind to the two G-protein coupled orexin receptors, OX1 and OX2, with orexin-A binding to both OX1 and OX2 with approximately equal affinity while orexin-B binds mainly to OX2 and is 5 times less potent at OX1.[11]
The orexins are strongly conserved peptides, found in all major classes of vertebrates.[12]
Function
The orexin system was initially suggested to be primarily involved in the stimulation of food intake, based on the finding that central administration of orexin-A and -B increased food intake. In addition, it stimulates wakefulness, regulates energy expenditure, and modulates visceral function.
Brown fat activation
Many studies support that the orexin neurons regulate brown adipose tissue (BAT) activity via the sympathetic nervous system to enhance energy expenditure.[13][14] Although orexin knockout mice was reported to show maldevelopment of brown adipose tissue (BAT) [15], subsequent report has shown normal development of BAT [16].
Wakefulness
Orexin seems to promote wakefulness. Recent studies indicate that a major role of the orexin system is to integrate metabolic, circadian and sleep debt influences to determine whether an animal should be asleep or awake and active. Orexin neurons strongly excite various brain nuclei with important roles in wakefulness including the dopamine, norepinephrine, histamine and acetylcholine systems[17][18] and appear to play an important role in stabilizing wakefulness and sleep.
The discovery that an orexin receptor mutation causes the sleep disorder canine narcolepsy[19] in Doberman Pinschers subsequently indicated a major role for this system in sleep regulation. Genetic knockout mice lacking the gene for orexin were also reported to exhibit narcolepsy.[20] Transitioning frequently and rapidly between sleep and wakefulness, these mice display many of the symptoms of narcolepsy. Researchers are using this animal model of narcolepsy to study the disease.[21] Narcolepsy results in excessive daytime sleepiness, inability to consolidate wakefulness in the day (and sleep at night), and cataplexy, which is the loss of muscle tone in response to strong, usually positive, emotions. Dogs that lack a functional receptor for orexin have narcolepsy, while animals and people lacking the orexin neuropeptide itself also have narcolepsy.
Central administration of orexin-A strongly promotes wakefulness, increases body temperature and locomotion, and elicits a strong increase in energy expenditure. Sleep deprivation also increases orexin-A transmission. The orexin system may thus be more important in the regulation of energy expenditure than food intake. In fact, orexin-deficient narcoleptic patients have increased obesity rather than decreased BMI, as would be expected if orexin were primarily an appetite stimulating peptide. Another indication that deficits of orexin cause narcolepsy is that depriving monkeys of sleep for 30–36 hours and then injecting them with the neurochemical alleviates the cognitive deficiencies normally seen with such amount of sleep loss.[22][23]
In humans, narcolepsy is associated with a specific variant of the human leukocyte antigen (HLA) complex.[24] Furthermore, genome-wide analysis shows that, in addition to the HLA variant, narcoleptic humans also exhibit a specific genetic mutation in the T-cell receptor alpha locus.[25] In conjunction, these genetic anomalies cause the immune system to attack and kill the critical orexin neurons. Hence the absence of orexin-producing neurons in narcoleptic humans may be the result of an autoimmune disorder.[26]
Food intake
Orexin increases the craving for food, and correlates with the function of the substances that promote its production. Orexin is also shown to increase meal size by suppressing inhibitory postingestive feedback.[27] However, some studies suggest that the stimulatory effects of orexin on feeding may be due to general arousal without necessarily increasing overall food intake.[28]
Review findings suggest that hyperglycemia that occurs in mice due to a habitual high-fat diet leads to a reduction in signalling by orexin receptor-2, and that orexin receptors may be a future therapeutic target.[29]
Leptin is a hormone produced by fat cells and acts as a long-term internal measure of energy state. Ghrelin is a short-term factor secreted by the stomach just before an expected meal, and strongly promotes food intake.
Orexin-producing cells have recently been shown to be inhibited by leptin (through the leptin receptor pathway), but are activated by ghrelin and hypoglycemia (glucose inhibits orexin production). Orexin, as of 2007, is claimed to be a very important link between metabolism and sleep regulation.[30][31] Such a relationship has been long suspected, based on the observation that long-term sleep deprivation in rodents dramatically increases food intake and energy metabolism, i.e., catabolism, with lethal consequences on a long-term basis. Sleep deprivation then leads to a lack of energy. In order to make up for this lack of energy, many people use high-carbohydrate and high-fat foods that ultimately can lead to poor health and weight gain. Other dietary nutrients, amino acids, also can activate orexin neurons, and they can suppress the glucose response of orexin neurons at physiological concentration, causing the energy balance that orexin maintains to be thrown off its normal cycle.[32]
Addiction
Preliminary research has been conducted that shows potential for orexin blockers in the treatment of cocaine, opioid, and alcohol addiction.[33][34][35] For example, lab rats given drugs which targeted the orexin system lost interest in alcohol despite being given free access in experiments.[36][37]
Studies of orexin involvement in nicotine addiction have had mixed results. For example, blocking the orexin-1 receptor with the selective orexin antagonist SB-334,867 reduced nicotine self-administration in rats and that smokers who suffered damage to the insula, a brain region that regulates cravings and contains orexin-1 receptors, lost the desire to smoke.[38] However, other studies in rats using the dual orexin receptor antagonist TCS 1102 have not found similar effects.[39]
Lipid metabolism
Orexin-A (OXA) has been recently demonstrated to have a direct effect on an aspect of lipid metabolism. OXA stimulates glucose uptake in 3T3-L1 adipocytes and that increased energy uptake is stored as lipids (triacylglycerol). OXA thus increases lipogenesis. It also inhibits lipolysis and stimulates the secretion of adiponectin. These effects are thought to be mostly conferred via the PI3K pathway because this pathway inhibitor (LY294002) completely blocks OXA effects in adipocytes.[40] The link between OXA and the lipid metabolism is new and currently under more research.
Mood
High levels of orexin-A have been associated with happiness in human subjects, while low levels have been associated with sadness.[41] The finding suggests that boosting levels of orexin-A could elevate mood in humans, being thus a possible future treatment for disorders like depression.
Orexin neurons
Neurotransmitters
Orexinergic neurons have been shown to be sensitive to inputs from Group III metabotropic glutamate receptors,[42] cannabinoid receptor 1 and CB1–OX1 receptor heterodimers,[43][44][45] adenosine A1 receptors,[46] muscarinic M3 receptors,[47] serotonin 5-HT1A receptors,[48] neuropeptide Y receptors,[49] cholecystokinin A receptors,[50] and catecholamines,[51][52] as well as to ghrelin, leptin, and glucose.[53] Orexinergic neurons themselves regulate release of acetylcholine,[54][55] serotonin, and noradrenaline.[56]
Orexinergic neurons can be differentiated into two groups based on connectivity and functionality. Orexinergic neurons in the lateral hypothalamic group are closely associated with reward related functions, such as conditioned place preference. These neurons preferentially innervate the ventral tegmental area and the ventromedial prefrontal cortex. In contrast to the lateral hypothalamic neurons, the perifornical-dorsal group of orexinergic neurons involved in functions related to arousal and autonomic response. These neurons project inter-hypothalamically, as well as to the brainstem, where the release of orexin modulates various autonomic processes.[57][58]
Clinical uses
The orexin/hypocretin system is the target of the insomnia medication suvorexant, which works by blocking both orexin receptors. Suvorexant has undergone three phase III trials and was approved in 2014 by the US Food and Drug Administration (FDA) after being denied approval the year before.[59] It is marketed as Belsomra.[60]
In 2016, the University of Texas Health Science Center registered a clinical trial for the use of suvorexant for people with cocaine dependence. They plan to measure cue reactivity, anxiety and stress.[61]
Other potential uses
Intranasal orexin is able to increase cognition in primates, especially under sleep deprived situations,[62] which may provide an opportunity for the treatment of excessive daytime sleepiness.[63]
A study has reported that transplantation of orexin neurons into the pontine reticular formation in rats is feasible, indicating the development of alternative therapeutic strategies in addition to pharmacological interventions to treat narcolepsy.[64]
References
- Davis JF, Choi DL, Benoit SC (2011). "24. Orexigenic Hypothalamic Peptides Behavior and Feeding - 24.5 Orexin". In Preedy VR, Watson RR, Martin CR (eds.). Handbook of Behavior, Food and Nutrition. Springer. pp. 361–2. ISBN 9780387922713.
- Stanford Center for Narcolepsy FAQ (retrieved 27-Mar-2012)
- Sutcliffe JG, de Lecea L (October 2000). "The hypocretins: excitatory neuromodulatory peptides for multiple homeostatic systems, including sleep and feeding". Journal of Neuroscience Research. 62 (2): 161–8. doi:10.1002/1097-4547(20001015)62:2<161::AID-JNR1>3.0.CO;2-1. PMID 11020209.
- Marcus JN, Elmquist JK (2006). "3. Orexin Projections and Localization of Orexin Receptors". In Nishino S, Sakurai T (eds.). The Orexin/Hypocretin System: Physiology and Pathophysiology. Springer. p. 195. ISBN 9781592599509.
- Boss C, Roch C (August 2015). "Recent trends in orexin research--2010 to 2015". Bioorganic & Medicinal Chemistry Letters. 25 (15): 2875–87. doi:10.1016/j.bmcl.2015.05.012. PMID 26045032.
- Sakurai T, Amemiya A, Ishii M, Matsuzaki I, Chemelli RM, Tanaka H, Williams SC, Richardson JA, Kozlowski GP, Wilson S, Arch JR, Buckingham RE, Haynes AC, Carr SA, Annan RS, McNulty DE, Liu WS, Terrett JA, Elshourbagy NA, Bergsma DJ, Yanagisawa M (February 1998). "Orexins and orexin receptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior". Cell. 92 (4): 573–85. doi:10.1016/S0092-8674(00)80949-6. PMID 9491897.
- de Lecea L, Kilduff TS, Peyron C, Gao X, Foye PE, Danielson PE, Fukuhara C, Battenberg EL, Gautvik VT, Bartlett FS, Frankel WN, van den Pol AN, Bloom FE, Gautvik KM, Sutcliffe JG (January 1998). "The hypocretins: hypothalamus-specific peptides with neuroexcitatory activity". Proceedings of the National Academy of Sciences of the United States of America. 95 (1): 322–7. doi:10.1073/pnas.95.1.322. PMC 18213. PMID 9419374.
- Gotter AL, Webber AL, Coleman PJ, Renger JJ, Winrow CJ (July 2012). "International Union of Basic and Clinical Pharmacology. LXXXVI. Orexin receptor function, nomenclature and pharmacology". Pharmacological Reviews. 64 (3): 389–420. doi:10.1124/pr.111.005546. PMID 22759794.
- Gautvik KM, de Lecea L, Gautvik VT, Danielson PE, Tranque P, Dopazo A, Bloom FE, Sutcliffe JG (August 1996). "Overview of the most prevalent hypothalamus-specific mRNAs, as identified by directional tag PCR subtraction". Proceedings of the National Academy of Sciences of the United States of America. 93 (16): 8733–8. doi:10.1073/pnas.93.16.8733. PMC 38742. PMID 8710940.
- Ebrahim IO, Howard RS, Kopelman MD, Sharief MK, Williams AJ (May 2002). "The hypocretin/orexin system". Journal of the Royal Society of Medicine. 95 (5): 227–30. doi:10.1177/014107680209500503. PMC 1279673. PMID 11983761.
- Langmead CJ, Jerman JC, Brough SJ, Scott C, Porter RA, Herdon HJ (January 2004). "Characterisation of the binding of [3H]-SB-674042, a novel nonpeptide antagonist, to the human orexin-1 receptor". British Journal of Pharmacology. 141 (2): 340–6. doi:10.1038/sj.bjp.0705610. PMC 1574197. PMID 14691055.
- Wong KK, Ng SY, Lee LT, Ng HK, Chow BK (April 2011). "Orexins and their receptors from fish to mammals: a comparative approach". General and Comparative Endocrinology. 171 (2): 124–30. doi:10.1016/j.ygcen.2011.01.001. PMID 21216246.
- Martins L, Seoane-Collazo P, Contreras C, González-García I, Martínez-Sánchez N, González F, Zalvide J, Gallego R, Diéguez C, Nogueiras R, Tena-Sempere M, López M (August 2016). "A Functional Link between AMPK and Orexin Mediates the Effect of BMP8B on Energy Balance". Cell Reports. 16 (8): 2231–42. doi:10.1016/j.celrep.2016.07.04. PMID 27524625.
- Tupone D, Madden CJ, Cano G, Morrison SF (November 2011). "An orexinergic projection from perifornical hypothalamus to raphe pallidus increases rat brown adipose tissue thermogenesis". Journal of Neuroscience. 31 (44): 15944–55. doi:10.1523/JNEUROSCI.3909-11.2011. PMID 22049437.
- Sellayah D, Bharaj P, Sikder D (October 2011). "Orexin is required for brown adipose tissue development, differentiation, and function". Cell Metabolism. 14 (4): 478–90. doi:10.1016/j.cmet.2011.08.010. PMID 21982708.
- Kakizaki M, Tsuneoka Y, Takase K, Kim SJ, Choi J, Ikkyu A, Abe M, Sakimura K, Yanagisawa M, Funato H (September 2019). "Differential Roles of Each Orexin Receptor Signaling in Obesity". iScience. 20. doi:10.1016/j.isci.2019.09.003. PMID 31546102.
- Sherin JE, Elmquist JK, Torrealba F, Saper CB (June 1998). "Innervation of histaminergic tuberomammillary neurons by GABAergic and galaninergic neurons in the ventrolateral preoptic nucleus of the rat". The Journal of Neuroscience. 18 (12): 4705–21. doi:10.1523/JNEUROSCI.18-12-04705.1998. PMID 9614245.
- Lu J, Bjorkum AA, Xu M, Gaus SE, Shiromani PJ, Saper CB (June 2002). "Selective activation of the extended ventrolateral preoptic nucleus during rapid eye movement sleep". The Journal of Neuroscience. 22 (11): 4568–76. PMID 12040064.
- Lin L, Faraco J, Li R, Kadotani H, Rogers W, Lin X, Qiu X, de Jong PJ, Nishino S, Mignot E (August 1999). "The sleep disorder canine narcolepsy is caused by a mutation in the hypocretin (orexin) receptor 2 gene". Cell. 98 (3): 365–76. doi:10.1016/S0092-8674(00)81965-0. PMID 10458611.
- Chemelli RM, Willie JT, Sinton CM, Elmquist JK, Scammell T, Lee C, Richardson JA, Williams SC, Xiong Y, Kisanuki Y, Fitch TE, Nakazato M, Hammer RE, Saper CB, Yanagisawa M (August 1999). "Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation". Cell. 98 (4): 437–51. doi:10.1016/S0092-8674(00)81973-X. PMID 10481909.
- Mochizuki T, Crocker A, McCormack S, Yanagisawa M, Sakurai T, Scammell TE (July 2004). "Behavioral state instability in orexin knock-out mice". The Journal of Neuroscience. 24 (28): 6291–300. doi:10.1523/JNEUROSCI.0586-04.2004. PMID 15254084.
- Alexis Madrigal (2007-12-28). "Snorting a Brain Chemical Could Replace Sleep". Wired News, Condé Nast. Retrieved 2008-02-05.
- Deadwyler SA, Porrino L, Siegel JM, Hampson RE (December 2007). "Systemic and nasal delivery of orexin-A (Hypocretin-1) reduces the effects of sleep deprivation on cognitive performance in nonhuman primates". The Journal of Neuroscience. 27 (52): 14239–47. doi:10.1523/JNEUROSCI.3878-07.2007. PMID 18160631.
- Klein J, Sato A (September 2000). "The HLA system. Second of two parts". The New England Journal of Medicine. 343 (11): 782–6. doi:10.1056/NEJM200009143431106. PMID 10984567.
- Hallmayer J, Faraco J, Lin L, Hesselson S, Winkelmann J, Kawashima M, Mayer G, Plazzi G, Nevsimalova S, Bourgin P, Hong SC, Hong SS, Honda Y, Honda M, Högl B, Longstreth WT, Montplaisir J, Kemlink D, Einen M, Chen J, Musone SL, Akana M, Miyagawa T, Duan J, Desautels A, Erhardt C, Hesla PE, Poli F, Frauscher B, Jeong JH, Lee SP, Ton TG, Kvale M, Kolesar L, Dobrovolná M, Nepom GT, Salomon D, Wichmann HE, Rouleau GA, Gieger C, Levinson DF, Gejman PV, Meitinger T, Young T, Peppard P, Tokunaga K, Kwok PY, Risch N, Mignot E (June 2009). "Narcolepsy is strongly associated with the T-cell receptor alpha locus". Nature Genetics. 41 (6): 708–11. doi:10.1038/ng.372. PMC 2803042. PMID 19412176.
- "Narcolepsy is an autoimmune disorder, Stanford researcher says". EurekAlert. American Association for the Advancement of Science. 2009-05-03. Retrieved 2009-05-31.
- Baird JP, Choe A, Loveland JL, Beck J, Mahoney CE, Lord JS, Grigg LA (March 2009). "Orexin-A hyperphagia: hindbrain participation in consummatory feeding responses". Endocrinology. 150 (3): 1202–16. doi:10.1210/en.2008-0293. PMC 2654731. PMID 19008313.
- Ida T, Nakahara K, Katayama T, Murakami N, Nakazato M (March 1999). "Effect of lateral cerebroventricular injection of the appetite-stimulating neuropeptide, orexin and neuropeptide Y, on the various behavioral activities of rats". Brain Research. 821 (2): 526–9. doi:10.1016/S0006-8993(99)01131-2. PMID 10064841.
- Tsuneki H, Wada T, Sasaoka T (March 2010). "Role of orexin in the regulation of glucose homeostasis". Acta Physiologica. 198 (3): 335–48. doi:10.1111/j.1748-1716.2009.02008.x. PMID 19489767.
- Brisbare-Roch C, Dingemanse J, Koberstein R, Hoever P, Aissaoui H, Flores S, Mueller C, Nayler O, van Gerven J, de Haas SL, Hess P, Qiu C, Buchmann S, Scherz M, Weller T, Fischli W, Clozel M, Jenck F (February 2007). "Promotion of sleep by targeting the orexin system in rats, dogs and humans". Nature Medicine. 13 (2): 150–5. doi:10.1038/nm1544. PMID 17259994.
- Sakurai T (March 2007). "The neural circuit of orexin (hypocretin): maintaining sleep and wakefulness". Nature Reviews. Neuroscience. 8 (3): 171–81. doi:10.1038/nrn2092. PMID 17299454.
- Inutsuka A, Yamanaka A (2013-03-06). "The physiological role of orexin/hypocretin neurons in the regulation of sleep/wakefulness and neuroendocrine functions". Frontiers in Endocrinology. 4 (18): 18. doi:10.3389/fendo.2013.00018. PMC 3589707. PMID 23508038.
- "Neurotransmitter Orexin Associated With Pleasure And Reward Pathways In The Brain". ScienceDaily. Retrieved 2018-05-08.
- Harris GC, Wimmer M, Aston-Jones G (September 2005). "A role for lateral hypothalamic orexin neurons in reward seeking". Nature. 437 (7058): 556–9. doi:10.1038/nature04071. PMID 16100511.
- Smith RJ, See RE, Aston-Jones G (August 2009). "Orexin/hypocretin signaling at the orexin 1 receptor regulates cue-elicited cocaine-seeking". The European Journal of Neuroscience. 30 (3): 493–503. doi:10.1111/j.1460-9568.2009.06844.x. PMC 2771107. PMID 19656173.
- Helen Puttick (2006-12-26). "Hope in fight against alcoholism". The Herald.
- Lawrence AJ, Cowen MS, Yang HJ, Chen F, Oldfield B (July 2006). "The orexin system regulates alcohol-seeking in rats". British Journal of Pharmacology. 148 (6): 752–9. doi:10.1038/sj.bjp.0706789. PMC 1617074. PMID 16751790.
- "Blocking A Neuropeptide Receptor Decreases Nicotine Addiction". ScienceDaily LLC. 2008-12-01. Retrieved 2009-02-11.
- Khoo SY, McNally GP, Clemens KJ (2017). "The dual orexin receptor antagonist TCS1102 does not affect reinstatement of nicotine-seeking". PLOS One. 12 (3): e0173967. doi:10.1371/journal.pone.0173967. PMC 5351999. PMID 28296947.
- Skrzypski M, T Le T, Kaczmarek P, Pruszynska-Oszmalek E, Pietrzak P, Szczepankiewicz D, Kolodziejski PA, Sassek M, Arafat A, Wiedenmann B, Nowak KW, Strowski MZ (July 2011). "Orexin A stimulates glucose uptake, lipid accumulation and adiponectin secretion from 3T3-L1 adipocytes and isolated primary rat adipocytes". Diabetologia. 54 (7): 1841–52. doi:10.1007/s00125-011-2152-2. PMID 21505958.
- Blouin AM, Fried I, Wilson CL, Staba RJ, Behnke EJ, Lam HA, Maidment NT, Karlsson KÆ, Lapierre JL, Siegel JM (2013). "Human hypocretin and melanin-concentrating hormone levels are linked to emotion and social interaction". Nature Communications. 4: 1547. doi:10.1038/ncomms2461. PMC 3595130. PMID 23462990. Lay summary – Science Daily.
- Acuna-Goycolea C, Li Y, Van Den Pol AN (March 2004). "Group III metabotropic glutamate receptors maintain tonic inhibition of excitatory synaptic input to hypocretin/orexin neurons". The Journal of Neuroscience. 24 (12): 3013–22. doi:10.1523/JNEUROSCI.5416-03.2004. PMID 15044540.
- Flores A, Maldonado R, Berrendero F (December 2013). "Cannabinoid-hypocretin cross-talk in the central nervous system: what we know so far". Frontiers in Neuroscience. 7: 256. doi:10.3389/fnins.2013.00256. PMC 3868890. PMID 24391536.
Direct CB1-HcrtR1 interaction was first proposed in 2003 (Hilairet et al., 2003). Indeed, a 100-fold increase in the potency of hypocretin-1 to activate the ERK signaling was observed when CB1 and HcrtR1 were co-expressed ... In this study, a higher potency of hypocretin-1 to regulate CB1-HcrtR1 heteromer compared with the HcrtR1-HcrtR1 homomer was reported (Ward et al., 2011b). These data provide unambiguous identification of CB1-HcrtR1 heteromerization, which has a substantial functional impact. ... The existence of a cross-talk between the hypocretinergic and endocannabinoid systems is strongly supported by their partially overlapping anatomical distribution and common role in several physiological and pathological processes. However, little is known about the mechanisms underlying this interaction.
• Figure 1: Schematic of brain CB1 expression and orexinergic neurons expressing OX1 or OX2
• Figure 2: Synaptic signaling mechanisms in cannabinoid and orexin systems
• Figure 3: Schematic of brain pathways involved in food intake - Thompson MD, Xhaard H, Sakurai T, Rainero I, Kukkonen JP (2014). "OX1 and OX2 orexin/hypocretin receptor pharmacogenetics". Frontiers in Neuroscience. 8: 57. doi:10.3389/fnins.2014.00057. PMC 4018553. PMID 24834023.
OX1–CB1 dimerization was suggested to strongly potentiate orexin receptor signaling, but a likely explanation for the signal potentiation is, instead, offered by the ability of OX1 receptor signaling to produce 2-arachidonoyl glycerol, a CB1 receptor ligand, and a subsequent co-signaling of the receptors (Haj-Dahmane and Shen, 2005; Turunen et al., 2012; Jäntti et al., 2013). However, this does not preclude dimerization.
- Jäntti MH, Mandrika I, Kukkonen JP (March 2014). "Human orexin/hypocretin receptors form constitutive homo- and heteromeric complexes with each other and with human CB1 cannabinoid receptors". Biochemical and Biophysical Research Communications. 445 (2): 486–90. doi:10.1016/j.bbrc.2014.02.026. PMID 24530395.
Orexin receptor subtypes readily formed homo- and hetero(di)mers, as suggested by significant BRET signals. CB1 receptors formed homodimers, and they also heterodimerized with both orexin receptors. ... In conclusion, orexin receptors have a significant propensity to make homo- and heterodi-/oligomeric complexes. However, it is unclear whether this affects their signaling. As orexin receptors efficiently signal via endocannabinoid production to CB1 receptors, dimerization could be an effective way of forming signal complexes with optimal cannabinoid concentrations available for cannabinoid receptors.
- Liu ZW, Gao XB (January 2007). "Adenosine inhibits activity of hypocretin/orexin neurons by the A1 receptor in the lateral hypothalamus: a possible sleep-promoting effect". Journal of Neurophysiology. 97 (1): 837–48. doi:10.1152/jn.00873.2006. PMC 1783688. PMID 17093123.
- Ohno K, Hondo M, Sakurai T (March 2008). "Cholinergic regulation of orexin/hypocretin neurons through M(3) muscarinic receptor in mice". Journal of Pharmacological Sciences. 106 (3): 485–91. doi:10.1254/jphs.FP0071986. PMID 18344611. Archived from the original on 2012-12-19.
- Muraki Y, Yamanaka A, Tsujino N, Kilduff TS, Goto K, Sakurai T (August 2004). "Serotonergic regulation of the orexin/hypocretin neurons through the 5-HT1A receptor". The Journal of Neuroscience. 24 (32): 7159–66. doi:10.1523/JNEUROSCI.1027-04.2004. PMID 15306649.
- Fu LY, Acuna-Goycolea C, van den Pol AN (October 2004). "Neuropeptide Y inhibits hypocretin/orexin neurons by multiple presynaptic and postsynaptic mechanisms: tonic depression of the hypothalamic arousal system". The Journal of Neuroscience. 24 (40): 8741–51. doi:10.1523/JNEUROSCI.2268-04.2004. PMID 15470140.
- Tsujino N, Yamanaka A, Ichiki K, Muraki Y, Kilduff TS, Yagami K, Takahashi S, Goto K, Sakurai T (August 2005). "Cholecystokinin activates orexin/hypocretin neurons through the cholecystokinin A receptor". The Journal of Neuroscience. 25 (32): 7459–69. doi:10.1523/JNEUROSCI.1193-05.2005. PMID 16093397.
- Li Y, van den Pol AN (January 2005). "Direct and indirect inhibition by catecholamines of hypocretin/orexin neurons". The Journal of Neuroscience. 25 (1): 173–83. doi:10.1523/JNEUROSCI.4015-04.2005. PMID 15634779.
- Yamanaka A, Muraki Y, Ichiki K, Tsujino N, Kilduff TS, Goto K, Sakurai T (July 2006). "Orexin neurons are directly and indirectly regulated by catecholamines in a complex manner". Journal of Neurophysiology. 96 (1): 284–98. CiteSeerX 10.1.1.320.260. doi:10.1152/jn.01361.2005. PMID 16611835.
- Ohno K, Sakurai T (January 2008). "Orexin neuronal circuitry: role in the regulation of sleep and wakefulness". Frontiers in Neuroendocrinology. 29 (1): 70–87. doi:10.1016/j.yfrne.2007.08.001. PMID 17910982.
- Bernard R, Lydic R, Baghdoyan HA (October 2003). "Hypocretin-1 causes G protein activation and increases ACh release in rat pons" (PDF). The European Journal of Neuroscience. 18 (7): 1775–85. doi:10.1046/j.1460-9568.2003.02905.x. PMID 14622212.
- Frederick-Duus D, Guyton MF, Fadel J (November 2007). "Food-elicited increases in cortical acetylcholine release require orexin transmission". Neuroscience. 149 (3): 499–507. doi:10.1016/j.neuroscience.2007.07.061. PMID 17928158.
- Soffin EM, Gill CH, Brough SJ, Jerman JC, Davies CH (June 2004). "Pharmacological characterisation of the orexin receptor subtype mediating postsynaptic excitation in the rat dorsal raphe nucleus". Neuropharmacology. 46 (8): 1168–76. doi:10.1016/j.neuropharm.2004.02.014. PMID 15111023.
- Aston-Jones G, Smith RJ, Sartor GC, Moorman DE, Massi L, Tahsili-Fahadan P, Richardson KA (February 2010). "Lateral hypothalamic orexin/hypocretin neurons: A role in reward-seeking and addiction". Brain Research. 1314: 74–90. doi:10.1016/j.brainres.2009.09.106. PMC 2819557. PMID 19815001.
- Grimaldi D, Silvani A, Benarroch EE, Cortelli P (January 2014). "Orexin/hypocretin system and autonomic control: new insights and clinical correlations". Neurology. 82 (3): 271–8. doi:10.1212/WNL.0000000000000045. PMID 24363130.
- Ventura, Jeff, ed. (2014-08-31). "FDA approves new type of sleep drug, Belsomra". Food and Drug Administration (FDA). Retrieved 2015-10-31.
- "BELSOMRA® (suvorexant) C-IV". Belsomra. Retrieved 2015-10-31.
- "Role of the Orexin Receptor System in Stress, Sleep and Cocaine Use (NCT02785406)". ClinicalTrials.gov. Retrieved 2017-07-08.
- Nixon JP, Mavanji V, Butterick TA, Billington CJ, Kotz CM, Teske JA (March 2015). "Sleep disorders, obesity, and aging: the role of orexin". Ageing Research Reviews. 20: 63–73. doi:10.1016/j.arr.2014.11.001. PMC 4467809. PMID 25462194.
- Billiard M (June 2008). "Narcolepsy: current treatment options and future approaches". Neuropsychiatric Disease and Treatment. 4 (3): 557–66. PMC 2526380. PMID 18830438.
- Arias-Carrión O, Murillo-Rodriguez E, Xu M, Blanco-Centurion C, Drucker-Colín R, Shiromani PJ (December 2004). "Transplantation of hypocretin neurons into the pontine reticular formation: preliminary results" (PDF). Sleep. 27 (8): 1465–70. doi:10.1093/sleep/27.8.1465. PMC 1201562. PMID 15683135. Archived from the original (PDF) on 2016-03-03.
External links
| Wikimedia Commons has media related to Orexin. |
- orexins at the US National Library of Medicine Medical Subject Headings (MeSH)
- Compare Different Sleep Aids, National Sleep Foundation
- Orexin receptor antagonists: A new class of sleeping pill, National Sleep Foundation