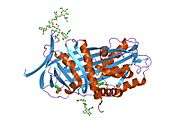
Alpha-1 antitrypsin
Alpha-1 antitrypsin or α1-antitrypsin (A1AT, A1A, or AAT) is a protein belonging to the serpin superfamily. It is encoded in humans by the SERPINA1 gene. A protease inhibitor, it is also known as alpha1–proteinase inhibitor (A1PI) or alpha1-antiproteinase (A1AP) because it inhibits various proteases (not just trypsin).[5] In older biomedical literature it was sometimes called serum trypsin inhibitor (STI, dated terminology), because its capability as a trypsin inhibitor was a salient feature of its early study. As a type of enzyme inhibitor, it protects tissues from enzymes of inflammatory cells, especially neutrophil elastase, and has a reference range in blood of 0.9–2.3 g/L (in the US the reference range is expressed as mg/dL or micromoles), but the concentration can rise manyfold upon acute inflammation.[6] When the blood contains inadequate amounts of A1AT or functionally defective A1AT (such as in alpha-1 antitrypsin deficiency), neutrophil elastase is excessively free to break down elastin, degrading the elasticity of the lungs, which results in respiratory complications, such as chronic obstructive pulmonary disease, in adults. Normally, A1AT leaves its site of origin, the liver, and joins the systemic circulation; defective A1AT can fail to do so, building up in the liver, which results in cirrhosis in either adults or children.


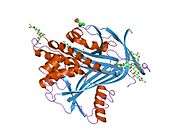
In addition to binding to neutrophil elastase released by inflammatory cells, A1AT also binds to elastase localized on the cell surface in which case elastase does not act as an enzyme, but instead acts to signal cells to undergo locomotion.[7]. Besides liver cells, A1PI is produced in bone marrow, by lymphocytic and monocytic cells in lymphoid tissue, and by the Paneth cells of the gut. [8]
Inactivation of A1AT by enzymes other than elastase due to inflammation/infection causes the migration of T cells to halt precisely at the site where the pathologic insult exists. This suggests a role for α1PI not only in locomotion of lymphocytes through tissue, but as a consequence of infection, a primary role as a sentinel in immune vigilance. [9]
A1AT is both an endogenous protease inhibitor and an exogenous one used as medication. The pharmaceutical form is purified from human donor blood and is sold under the nonproprietary name alpha1–proteinase inhibitor (human) and under various trade names (including Aralast NP, Glassia, Prolastin, Prolastin-C, and Zemaira). Recombinant versions are also available but are currently used in medical research more than as medication.
Function
A1AT is a 52-kDa serpin and, in medicine, it is considered the most prominent serpin; the terms α1-antitrypsin and protease inhibitor (Pi) are often used interchangeably.
Most serpins inactivate enzymes by binding to them covalently. These enzymes are released locally in relatively low concentrations where they are immediately cleared by proteins such as A1AT. In the acute phase reaction, a further elevation is required to "limit" the damage caused by activated neutrophil granulocytes and their enzyme elastase, which breaks down the connective tissue fiber elastin.
Besides limiting elastase activity to limit tissue degradation, A1PI also acts to induce locomotion of lymphocytes through tissue including immature T cells through the thymus where immature T cells mature to become immunocompetent T cells that are released into tissue to elevate immune responsiveness. [10]
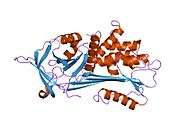
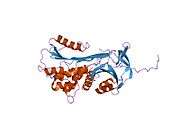
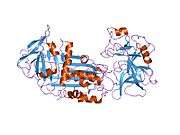
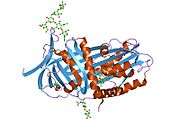
Like all serine protease inhibitors, A1AT has a characteristic secondary structure of beta sheets and alpha helices. Mutations in these areas can lead to non-functional proteins that can polymerise and accumulate in the liver (infantile hepatic cirrhosis).
Role in disease
.png)
Disorders of this protein include alpha-1 antitrypsin deficiency, an autosomal codominant hereditary disorder in which a deficiency of alpha-1 antitrypsin leads to a chronic uninhibited tissue breakdown. This causes the degradation especially of lung tissue and eventually leads to characteristic manifestations of pulmonary emphysema.[11] Evidence has shown[12] that cigarette smoke can result in oxidation of methionine 358 of α1-antitrypsin (382 in the pre-processed form containing the 24 amino acid signal peptide), a residue essential for binding elastase; this is thought to be one of the primary mechanisms by which cigarette smoking (or second-hand smoke) can lead to emphysema. Because A1AT is expressed in the liver, certain mutations in the gene encoding the protein can cause misfolding and impaired secretion, which can lead to liver cirrhosis.
An extremely rare form of Pi, termed PiPittsburgh, functions as an antithrombin (a related serpin), due to a mutation (Met358Arg). One person with this mutation has been reported to have died of a bleeding diathesis.[13]
A liver biopsy will show abundant PAS-positive globules within periportal hepatocytes.
Patients with rheumatoid arthritis (RA) have been found to make autoantibodies toward the carbamylated form of A1AT in the synovial fluid. This suggests that A1AT may play an anti-inflammatory or tissue-protecting role outside the lungs. These antibodies are associated with a more severe disease course, can be observed years before disease onset, and may predict the development of RA in arthralgia patients. Consequently, carbamylated A1AT is currently being developed as an antigenic biomarker for RA.[14]
Nomenclature
The protein was initially named "antitrypsin" because of its ability to bind and irreversibly inactivate the enzyme trypsin in vitro covalently. Trypsin, a type of peptidase, is a digestive enzyme active in the duodenum and elsewhere.
The term alpha-1 refers to the protein's behavior on protein electrophoresis. On electrophoresis, the protein component of the blood is separated by electric current. There are several clusters, the first being albumin, the second being the alpha, the third beta and the fourth gamma (immunoglobulins). The non-albumin proteins are referred to as globulins.
The alpha region can be further divided into two sub-regions, termed "1" and "2". Alpha-1 antitrypsin is the main protein of the alpha-globulin 1 region.
Another name used is alpha-1 proteinase inhibitor (α1-PI).
Genetics
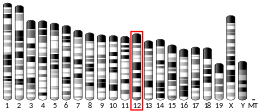
The gene is located on the long arm of chromosome 14 (14q32.1).
Over 100 different variants of α1-antitrypsin have been described in various populations. North-Western Europeans are most at risk for carrying one of the most common mutant forms of A1AT, the Z mutation (Glu342Lys on M1A, rs28929474).
Biochemical properties
A1AT is a single-chain glycoprotein consisting of 394 amino acids in the mature form and exhibits many glycoforms. The three N-linked glycosylations sites are mainly equipped with so-called diantennary N-glycans. However, one particular site shows a considerable amount of heterogeneity since tri- and even tetraantennary N-glycans can be attached to the Asparagine 107 (UniProtKB amino acid nomenclature). These glycans carry different amounts of negatively charged sialic acids; this causes the heterogeneity observed on normal A1AT when analysed by isoelectric focusing. Also, the fucosylated triantennary N-glycans were shown to have the fucose as part of a so-called Sialyl Lewis x epitope, which could confer this protein particular protein-cell recognition properties. The single cysteine residue of A1AT in position 256 (UniProtKB nomenclature) is found to be covalently linked to a free single cysteine by a disulfide bridge.[15]
Analysis
The level of A1AT in serum is most often determined by adding an antibody that binds to A1AT, then using turbidimetry to measure how much A1AT is present. Other detection methods include the use of enzyme-linked-immuno-sorbent-assays and radial immunodiffusion.
Different analytical methods are used to determine A1AT phenotype. As protein electrophoresis is imprecise, the A1AT phenotype is analysed by isoelectric focusing (IEF) in the pH range 4.5-5.5, where the protein migrates in a gel according to its isoelectric point or charge in a pH gradient.
Normal A1AT is termed M, as it migrates toward the center of such an IEF gel. Other variants are less functional and are termed A-L and N-Z, dependent on whether they run proximal or distal to the M band. The presence of deviant bands on IEF can signify the presence of alpha-1 antitrypsin deficiency. Since the number of identified mutations has exceeded the number of letters in the alphabet, subscripts have been added to most recent discoveries in this area, as in the Pittsburgh mutation described above.
As every person has two copies of the A1AT gene, a heterozygote with two different copies of the gene may have two different bands showing on electrofocusing, although heterozygote with one null mutant that abolishes expression of the gene will only show one band.
In blood test results, the IEF results are notated as in PiMM, where Pi stands for protease inhibitor and "MM" is the banding pattern of that patient.
Alpha-1 antitrypsin levels in the blood depend on the genotype. Some mutant forms fail to fold properly and are, thus, targeted for destruction in the proteasome, whereas others have a tendency to polymerise, being retained in the endoplasmic reticulum. The serum levels of some of the common genotypes are:
- PiMM: 100% (normal)
- PiMS: 80% of normal serum level of A1AT
- PiSS: 60% of normal serum level of A1AT
- PiMZ: 60% of normal serum level of A1AT
- PiSZ: 40% of normal serum level of A1AT
- PiZZ: 10-15% (severe alpha-1 antitrypsin deficiency)
- PiZ is caused by a glutamate to lysine mutation at position 342 (366 in pre-processed form)
- PiS is caused by a glutamate to valine mutation at position 264 (288 in pre-processed form)
Other rarer forms have been described; in all, there are over 80 variants.
Medical use
 | |
| Clinical data | |
|---|---|
| Routes of administration | intravenous |
| ATC code | |
| Identifiers | |
IUPAC name
| |
| CAS Number | |
| DrugBank | |
| ChemSpider |
|
| ECHA InfoCard | 100.029.919 |
| Chemical and physical data | |
| Formula | C2001H3130N514O601S10 |
| Molar mass | 44324.5 g/mol g·mol−1 |
| (verify) | |
Alpha-1 antitrypsin concentrates are prepared from the blood plasma of blood donors. The US FDA has approved the use of four alpha-1 antitrypsin products derived from a human plasma: Prolastin, Zemaira, Glassia, and Aralast. These products for intravenous augmentation A1AT therapy can cost up to $100,000 per year per patient.[16] They are administered intravenously at a dose of 60 mg/kg once a week; higher doses do not provided additional benefit although they can be used in anticipation of an interruption of weekly administration, such as for a vacation.[17]
Aerosolized-augmented A1AT therapy is under study. This involves inhaling purified human A1AT into the lungs and trapping the A1AT into the lower respiratory tract. However, inhaled A1AT may not reach the elastin fibers in the lung where elastase injury occurs. Further study is currently underway. Recombinant alpha-1 antitrypsin is not yet available for use as a medication but is under development.
History
Axelsson and Laurell first investigated the possibility of allelic variants of A1AT leading to disease in 1965.[18]
See also
- Alpha 1-antichymotrypsin, another serpin that is analogous for protecting the body from excessive effects of its own inflammatory proteases
References
- ENSG00000277377 GRCh38: Ensembl release 89: ENSG00000197249, ENSG00000277377 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000071177 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Gettins PG (December 2002). "Serpin structure, mechanism, and function". Chemical Reviews. 102 (12): 4751–804. doi:10.1021/cr010170. PMID 12475206.
- Kushner, Mackiewicz A (1993). The acute phase response: an overview. Acute-phase glycoproteins: molecular biology, biochemistry and clinical applications. CRC Press. pp. 3–19.
- Guttman O, Baranovski BM, Schuster R, Kaner Z, Freixo-Lima GS, Bahar N, Mizrahi MI, Brami I, Ochayon DE, Lewis EC (February 2015). "Acute-phase protein α1-anti-trypsin: diverting injurious innate and adaptive immune responses from non-authentic threats". Clinical & Experimental Immunology. 179 (2): 161–172. doi:10.1111/cei.12476. PMC 4298394. PMID 25351931.
- Winkler IG, Hendy J, Coughlin P, Horvath A, Lévesque JP (April 2005). "Serine protease inhibitors serpina1 and serpina3 are down-regulated in bone marrow during hematopoietic progenitor mobilization". The Journal of Experimental Medicine. 201 (7): 1077–88. doi:10.1084/jem.20042299. PMC 2213124. PMID 15795238.
- Richler R, Forssmann W, Henschler R (June 2017). "Current developments in mobilization of hematopoietic stem and progenitor cells and their interaction with nicehes in bone marrow". Transfus Med Hemother. 44 (3). doi:10.1159/000477262. PMC 5473067. PMID 28626366.
- Lapidot T, Petit I (September 2012). "Current understanding of stem cell mobilization: the roles of chemokines, proteolytic enzymes, adhesion molecules, cytokines, and stromal cells". Exp Hematol. 30 (9): 973–981. PMID 12225788.
- DeMeo DL, Silverman EK (March 2004). "Alpha1-antitrypsin deficiency. 2: genetic aspects of alpha(1)-antitrypsin deficiency: phenotypes and genetic modifiers of emphysema risk". Thorax. 59 (3): 259–64. doi:10.1136/thx.2003.006502. PMC 1746953. PMID 14985567.
- Taggart C, Cervantes-Laurean D, Kim G, McElvaney NG, Wehr N, Moss J, Levine RL (September 2000). "Oxidation of either methionine 351 or methionine 358 in alpha 1-antitrypsin causes loss of anti-neutrophil elastase activity". The Journal of Biological Chemistry. 275 (35): 27258–65. doi:10.1074/jbc.M004850200. PMID 10867014.
- Owen MC, Brennan SO, Lewis JH, Carrell RW (September 1983). "Mutation of antitrypsin to antithrombin. alpha 1-antitrypsin Pittsburgh (358 Met leads to Arg), a fatal bleeding disorder". The New England Journal of Medicine. 309 (12): 694–8. doi:10.1056/NEJM198309223091203. PMID 6604220.
- Verheul MK, Yee A, Seaman A, Janssen GM, van Veelen PA, Drijfhout JW, Toes RE, Mahler M, Trouw LA (June 2017). "Identification of carbamylated alpha 1 anti-trypsin (A1AT) as an antigenic target of anti-CarP antibodies in patients with rheumatoid arthritis". Journal of Autoimmunity. 80: 77–84. doi:10.1016/j.jaut.2017.02.008. PMID 28291659.
- Kolarich D, Weber A, Turecek PL, Schwarz HP, Altmann F (June 2006). "Comprehensive glyco-proteomic analysis of human alpha1-antitrypsin and its charge isoforms". Proteomics. 6 (11): 3369–80. doi:10.1002/pmic.200500751. PMID 16622833.
- Alkins SA, O'Malley P (March 2000). "Should health-care systems pay for replacement therapy in patients with alpha(1)-antitrypsin deficiency? A critical review and cost-effectiveness analysis". Chest. 117 (3): 875–80. doi:10.1378/chest.117.3.875. PMID 10713018.
- Brantly, Mark L.; Lascano, Jorge E.; Shahmohammadi, Abbas (2018-11-28). "Intravenous Alpha-1 Antitrypsin Therapy for Alpha-1 Antitrypsin Deficiency: The Current State of the Evidence". Chronic Obstructive Pulmonary Diseases (Miami, Fla.). 6 (1): 100–114. doi:10.15326/jcopdf.6.1.2017.0185. PMC 6373587. PMID 30775428.
- Axelsson U, Laurell CB (November 1965). "Hereditary variants of serum alpha-1-antitrypsin". American Journal of Human Genetics. 17 (6): 466–72. PMC 1932630. PMID 4158556.
Further reading
- Kalsheker N (April 1989). "Alpha 1-antitrypsin: structure, function and molecular biology of the gene". Bioscience Reports. 9 (2): 129–38. doi:10.1007/BF01115992. PMID 2669992.
- Crystal RG (December 1989). "The alpha 1-antitrypsin gene and its deficiency states". Trends in Genetics. 5 (12): 411–7. doi:10.1016/0168-9525(89)90200-X. PMID 2696185.
- Carrell RW, Jeppsson JO, Laurell CB, Brennan SO, Owen MC, Vaughan L, Boswell DR (July 1982). "Structure and variation of human alpha 1-antitrypsin". Nature. 298 (5872): 329–34. doi:10.1038/298329a0. PMID 7045697.
- Elliott PR, Abrahams JP, Lomas DA (January 1998). "Wild-type alpha 1-antitrypsin is in the canonical inhibitory conformation". Journal of Molecular Biology. 275 (3): 419–25. doi:10.1006/jmbi.1997.1458. PMID 9466920.
- Miyamoto Y, Akaike T, Maeda H (March 2000). "S-nitrosylated human alpha(1)-protease inhibitor". Biochimica et Biophysica Acta. 1477 (1–2): 90–7. doi:10.1016/S0167-4838(99)00264-2. PMID 10708851.
- Coakley RJ, Taggart C, O'Neill S, McElvaney NG (January 2001). "Alpha1-antitrypsin deficiency: biological answers to clinical questions". The American Journal of the Medical Sciences. 321 (1): 33–41. doi:10.1097/00000441-200101000-00006. PMID 11202478.
- Lomas DA, Lourbakos A, Cumming SA, Belorgey D (April 2002). "Hypersensitive mousetraps, alpha1-antitrypsin deficiency and dementia". Biochemical Society Transactions. 30 (2): 89–92. doi:10.1042/BST0300089. PMID 12023831.
- Kalsheker N, Morley S, Morgan K (April 2002). "Gene regulation of the serine proteinase inhibitors alpha1-antitrypsin and alpha1-antichymotrypsin". Biochemical Society Transactions. 30 (2): 93–8. doi:10.1042/BST0300093. PMID 12023832.
- Perlmutter DH (December 2002). "Liver injury in alpha1-antitrypsin deficiency: an aggregated protein induces mitochondrial injury". The Journal of Clinical Investigation. 110 (11): 1579–83. doi:10.1172/JCI16787. PMC 151639. PMID 12464659.
- Lomas DA, Mahadeva R (December 2002). "Alpha1-antitrypsin polymerization and the serpinopathies: pathobiology and prospects for therapy". The Journal of Clinical Investigation. 110 (11): 1585–90. doi:10.1172/JCI16782. PMC 151637. PMID 12464660.
- Lisowska-Myjak B (February 2005). "AAT as a diagnostic tool". Clinica Chimica Acta; International Journal of Clinical Chemistry. 352 (1–2): 1–13. doi:10.1016/j.cccn.2004.03.012. PMID 15653097.
- Lomas DA (2005). "Molecular mousetraps, alpha1-antitrypsin deficiency and the serpinopathies". Clinical Medicine. 5 (3): 249–57. doi:10.7861/clinmedicine.5-3-249. PMC 4952210. PMID 16011217.
- Rudnick DA, Perlmutter DH (September 2005). "Alpha-1-antitrypsin deficiency: a new paradigm for hepatocellular carcinoma in genetic liver disease". Hepatology. 42 (3): 514–21. doi:10.1002/hep.20815. PMID 16044402.
External links
- The MEROPS online database for peptidases and their inhibitors: I04.001
- Proteopedia: Alpha-1-antitrypsin
- Alpha-1 antitrypsin at Lab Tests Online
- Alpha-1 Disease Management Program at AlphaNet
- Frequently Asked Questions (FAQ) about Alpha-1 from AlphaNet
- Human SERPINA1 genome location and SERPINA1 gene details page in the UCSC Genome Browser.
- Overview of all the structural information available in the PDB for UniProt: P01009 (Alpha-1-antitrypsin) at the PDBe-KB.
PDB gallery | |
|---|---|
|