ஈரலழற்சி பி தீநுண்மம்
எட்.பி.வி என சுருக்கி அழைக்கப்படும் ஈரலழற்சி பி தீநுண்மம் என்பது ஆர்த்தோஹெபாட்னாவைரஸ் (Orthohepadnavirus) பேரினத்தைச் சேர்ந்த ஒரு இனமாகும். இதே போன்று, இந்த பேரினமானது ஹெபாட்னாவிரிடே (Hepadnaviridae) குடும்பத்தைச் சேர்ந்த தீநுண்மங்களின் ஒரு பகுதியாகும்.[1] இந்தத் தீநுண்மம் ஈரலழற்சி பி நோய்க்குக் காரணமாகும்.[2]
| Hepatitis B virus | |
|---|---|
 | |
| Micrograph showing hepatitis B viruses | |
| Virus classification | |
| குழு: | Group VII (dsDNA-RT) |
| வரிசை: | Unassigned |
| குடும்பம்: | Hepadnaviridae |
| பேரினம்: | Orthohepadnavirus |
| இனம்: | Hepatitis B virus |
நோய்
ஈரலழற்சி பி -க்குக் காரணமாவதுடன், எச்.பி.வி இன் நோய்த்தொற்று கல்லீரலில் இழைநார் வளர்ச்சி (cirrhosis) மற்றும் ஈரல்செல்லியப் புற்றுநோய் (hepatocellular carcinoma) ஆகியவற்றுக்கு வழிவகுக்கலாம்.[3]
மேலும் இது கணையச்சிரைப் புற்றுநோயின் இடர்பாட்டையும் அதிகரிக்கலாம் எனக் கூறப்படுகிறது.[2]
நோய் வகைப்பாடு
ஈரலழற்சி பி தீநுண்மம் ஆர்த்தோஹெபாட்னாவைரஸ் இனமாக வகைப்படுத்தப்படுகிறது. அது கிரவுண்ட் ஸ்க்விரில் ஹெபாடைடிஸ் வைரஸ் (Ground squirrel hepatitis virus), உட்சக் ஹெபாடிடிஸ் வைரஸ் (Woodchuck hepatitis virus) மற்றும் ஊல்லி மங்கி ஹெபாடைடிஸ் பி வைரஸ் (Woolly monkey hepatitis B virus) ஆகிய மூன்று மற்ற இனங்களையும் கொண்டிருக்கிறது. இந்தப் பேரினம் ஹெபாட்னாவிரிடே குடும்பத்தின் ஒரு பகுதியாக வகைப்படுத்தப்படுகிறது. அது அவிஹெபாட்னாவைரஸ் (Avihepadnavirus) மற்றும் இன்னும் பெயரிடப்படாத ஒன்று ஆகிய இரண்டு மற்ற இனங்களைக் கொண்டிருக்கிறது. தீநுண்மங்களின் இந்தக் குடும்பமானது நச்சுயிரி சார்ந்த ஒரு வரிசையில் வகைப்படுத்தப்படவில்லை.[4] ஈரலழற்சி பி போன்ற தீநுண்மங்கள் அனைத்து பழைய உலக வாலில்லாக் குரங்குகளில் (உர்ராங்குட்டான், கிப்பன்கள், கொரில்லாக்கள் மற்றும் சிம்பன்சிக்கள்) மற்றும் நவீன உலக கம்பளிக் குரங்கு போன்றவற்றில் கண்டுபிடிக்கப்பட்டுள்ளன. இக்கண்டுபிடிப்பானது உயர் விலங்கினங்களில் இந்த தீநுண்மத்துக்கான தொன்மையான துவக்கம் இருப்பதை அறிவுறுத்துகிறது.
இந்தத் தீநுண்மம் அதன் உறைப் புரதங்களில் இருக்கின்ற எதிர்செனிக் எபிடோப்புகளைப் பொறுத்து நான்கு முக்கிய குருதி நுண்ணுயிர் வகைகளாக (adr, adw, ayr, ayw) பிரிக்கப்படுகிறது. மேலும் மரபுத்தொகுதியின் ஒட்டுமொத்த நியூக்ளியோட்டைடு வரிசைமுறை மாறுபாடுகள் சார்ந்து எட்டு மரபணு அமைப்புகளாகப் (A-H) பிரிக்கப்படுகிறது. இந்த மரபணு அமைப்புகள் தனிப்பட்ட புவியியலுக்குரிய பரவலைக் கொண்டிருக்கின்றன. மேலும் அவை தீநுண்மத்தின் பரிணாமம் மற்றும் பரவுதலைக் கண்டுபிடித்தலில் பயன்படுகின்றன. மரபணு அமைப்புகளுக்கு இடையில் உள்ள மாறுபாடுகள் நோயின் தீவிரத்தன்மை, சிக்கல் நிலையின் செயல்பாடு மற்றும் நிகழும் வாய்ப்பு மற்றும் சிகிச்சை மற்றும் கூடுமாயின் தடுப்புமருந்து ஆகியவற்றுக்கான பிரதிவினை ஆகியவற்றைப் பாதிக்கின்றன.[5][6]
உருவியல்
கட்டமைப்பு
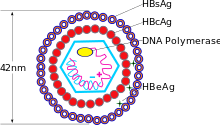
ஈரலழற்சி பி தீநுண்மம் (எச்.பி.வி) ஹெபாட்னாவைரஸ் குடும்பத்தின் (Hepadnavirus family) உறுப்பினராகும்.[7] தீநுண்மத் துகள் (முதிர்ந்த நச்சுயிரி (virion)) வெளிப்புற லிப்பிடு உறை மற்றும் புரதத்தினால் உருவாக்கப்பட்ட ஐகோசாஹேட்ரல் (icosahedral) அதிநுண்ணுயிர் அமைப்பு அடிப்படை ஆகியவற்றைக் கொண்டிருக்கிறது. இதில் அதிநுண்ணுயிர் அமைப்பு நச்சுயிரி சார்ந்த டி.என்.ஏ ஐயும், பின்திரும்பும் தீநுண்மங்கள் போன்ற நேர்மாறு படியெடுத்தல் நடவடிக்கையைக் கொண்டிருக்கும் டி.என்.ஏ. பாலிமரேசையும் உள்ளடக்கும்.[8] வெளிப்புற உறையில் பதியப்பட்ட புரதங்கள் உள்ளன. அவை எளிதில் பாதிக்கப்படக்கூடிய செல்களின் நச்சுயிரி சார்ந்த பிணைப்பு மற்றும் அத்தகைய செல்களில் நுழைதல் ஆகியவற்றுடன் தொடர்புடையனவாக உள்ளன. இந்தத் தீநுண்மம் 42 நேமீ முதிர்ந்த நச்சுயிரியுடன் கூடிய மிகவும் சிறிய உறையிடப்பட்ட விலங்கு தீநுண்மங்களில் ஒன்றாகும். ஆனால் இழை வடிவமுடைய மற்றும் கோளவுருவான அங்கங்களைக் கொண்டு மைய அமைப்பைக் கொண்டிராத வடிவம் மாறும் வடிவங்களும் உள்ளன. இந்தத் துகள்கள் தொற்றும் தன்மை கொண்டவை அல்ல. மேலும் இவை முதிர்ந்த நச்சுயிரியின் புறப்பரப்பின் பகுதியாக உருவாகும் லிப்பிடு மற்றும் புரதத்தால் உருவாகின்றன. இது புறப்பரப்பு எதிர்ச்செனி (HBsAg) என அழைக்கப்படுகிறது. மேலும் இது தீநுண்மத்தின் வாழ்க்கைச் சுழற்சி சமயத்தில் மிகுதியாக உருவாக்கப்படும்.[9]
மரபுத்தொகுதி
.
அளவு
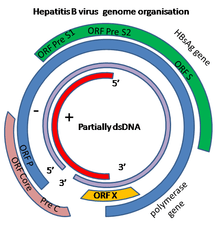
எச்.பி.வி இன் மரபுத்தொகுதி வட்டமான டி.என்.ஏ. கொண்டு உருவாகிறது. ஆனால் டி.என்.ஏ முழுமையாக இரட்டை-இழையிடப்பட்டது அல்ல என்பதால் இது அசாதாரணமானது. முழு நீள இழையின் ஒரு முனை நச்சுயிரி சார்ந்த டி.என்.ஏ. பாலிமரேஸ் உடன் இணைக்கப்பட்டிருக்கிறது. மரபுத்தொகுதியானது (முழு நீள இழைகளுக்கு) 3020-3320 நியூக்ளியோட்டைடுகள் நீளமாகவும், (குறை நீள இழைகளுக்கு) 1700-2800 நியூக்ளியோட்டைடுகள் நீளமாகவும் உள்ளது.[12]
குறியேற்றம்
எதிர்மறை-உணர்வு (குறியீடல்லாத) நச்சுயிரி சார்ந்த mRNA க்கு பூர்த்தி செய்வதாக இருக்கிறது. நச்சுயிரி சார்ந்த டி.என்.ஏவானது கலத்தில் தொற்று ஏற்பட்டதும் விரைவில் அணு உட்கருவில் காணப்படுகிறது. பகுதியளவு இரட்டை-இழையிடப்பட்ட டி.என்.ஏவானது (+) உணர்வு இழையின் நிறைவு, (-) உணர்வு இழையில் இருந்து புரத மூலக்கூறுகளை நீக்குதல் மற்றும் (+) உணர்வு இழையில் இருந்து ஆர்.என்.ஏ இன் குறுகிய வரிசை ஆகியவற்றின் மூலமாக முழு இரட்டை-இழையிடப்பட்டதை உருவாக்குகிறது. குறியீடல்லாத அடித்தளங்கள் (-)உணர்வு இழையின் முனைகளில் இருந்து நீக்கப்படுகின்றன பின்னர் முனைகள் மீண்டும் இணைக்கப்படுகின்றன.
C, X, P மற்றும் S என்று அழைக்கப்படும் நான்கு அறியப்பட்ட மரபணுக்கள் மரபுத்தொகுதி மூலமாக குறியீடு செய்யப்படுகின்றன. அடிப்படை புரதமானது C (HBcAg) மரபணு மூலமாக குறியீடாக்கப்படுகிறது. மேலும் அதன் ஆரம்பக் குறியீட்டு மொழி முன்-அடிப்படைப் புரதம் உருவாக்கப்படுவதில் இருந்து ஆற்றெதிர் உள்-சட்ட AUG ஆரம்பக் குறியீட்டு மொழி மூலமாக முன்செல்கிறது. HBeAg ஆனது முன்-அடிப்படைப் புரதத்தின் புரதச் சிதைப்பு செயல்பாடு மூலமாக உருவாகிறது. டி.என்.ஏ. பாலிமரேசு P மரபணு மூலமாக குறியீடாக்கப்படுகிறது. S மரபணு புறப்பரப்பு எதிர்ச்செனிக்காக (HBsAg) குறியிடப்படும் மரபணுவாக இருக்கிறது. HBsAg மரபணு ஒரு நீண்ட திறந்த படிக்கும் சட்டத்தைக் கொண்டிருக்கிறது. ஆனால் மூன்று உள்-சட்ட "ஆரம்ப" (ATG) குறியீட்டு மொழிகளைக் கொண்டிருக்கிறது. அது மரபணுவை pre-S1, pre-S2 மற்றும் S ஆகிய மூன்று பகுதிகளாகப் பிரிக்கிறது. பல ஆரம்பக் குறியீட்டு மொழிகள் இருப்பதன் காரணமாக பெரியது, நடுத்தரம் மற்றும் சிறியது (pre-S1 + pre-S2 + S, pre-S2 + S அல்லது S) என்று அழைக்கப்படும் மூன்று மாறுபட்ட அளவுகளில் பல பெப்டைடுகள் உருவாகின்றன.[13] மரபணு X மூலமாகக் குறியிடப்படும் புரதத்தின் செயல்பாடு முழுமையாக அறியப்படவில்லை.[14]
மரபணு அமைப்புகள்
A முதல் H வரையில் பெயரிடப்பட்ட எட்டு அறியப்பட மரபணு அமைப்புகள் இருக்கின்றன.[5] சாத்தியமுள்ள புதிய "I" மரபணு அமைப்பு விவரிக்கப்பட வேண்டியிருக்கிறது.[15] ஆனால் இந்தக் குறிமுறையின் ஏற்றுக்கொள்ளல் உலகளாவியதாக இல்லை.[16] மாறுபட்ட மரபணு அமைப்புகள் மாறுபட்ட வழிகளில் சிகிச்சைக்குப் பதிலளிக்கலாம்.[17][18]
குறைந்த 8% வரிசைமுறைகள் மூலமாக மரபணு அமைப்புகள் மாறுபடுகின்றன. மேலும் மாறுபட்ட புவியியலுக்குரிய விநியோகங்களைக் கொண்டிருக்கின்றன. மேலும் இது மானிடவியல் சார் வரலாற்றுடன் தொடர்புடையதாக இருக்கின்றது. 14% மற்ற மரபணுத் தொகுதிகளில் இருந்து பிரிந்து செல்லும் வகை F ஆனது அறியப்பட்டவைகளில் மிகவும் வேறுபட்ட வகையாகும். வகை A ஆனது ஐரோப்பா, ஆப்பிரிக்கா மற்றும் பிலிப்பைன்ஸ் உட்பட தென்-கிழக்கு ஆசியா ஆகிய நாடுகளில் பரவியிருக்கிறது. வகை B மற்றும் C ஆசியாவில் ஆதிக்கம் செலுத்துவதாக இருக்கிறது; வகை D ஆனது மத்தியத்தரைப் பகுதிகளான மத்திய கிழக்கு மற்றும் இந்தியாவில் பொதுவானதாக இருக்கிறது. வகை E ஆனது உப-சகாரா ஆப்பிரிக்காவில் குறிப்பிட்ட இடத்தில் இருக்கிறது. வகை F (அல்லது H) ஆனது மத்திய மற்றும் தென் அமெரிக்காவில் எல்லைக்குட்பட்டதாக இருக்கிறது. வகை G பிரான்சு மற்றும் ஜெர்மனியில் காணப்படுகின்றது. A, D மற்றும் F ஆகிய மரபணு அமைப்புகள் பிரேசிலில் ஆதிக்கம் செலுத்துவதாக இருக்கின்றன. மேலும் அமெரிக்க ஒன்றியத்தில் இனம் சார்ந்த அளவுகளுடன் அனைத்து மரபணு அமைப்புகளும் இருக்கின்றன.
E மற்றும் F திரிபுகள் முறையே ஆப்பிரிக்காவின் பழங்குடி மக்கள் மற்றும் நவீன உலகம் ஆகியவற்றில் தொடங்கியது.
மரபணு அமைப்புகளுக்குள் 24 உபவகைகள் விவரிக்கப்பட்டிருக்கின்றன. இவை 4-8% மரபுத்தொகுதி மூலமாக வேறுபடுத்தப்படுகிறது.
வகை A ஆனது ஆப்பிரிக்கா/ஆசியா மற்றும் பிலிப்பைன் ஆகிய பகுதிகளில் Aa (A1) மற்றும் ஐரோப்பா/அமெரிக்க ஒன்றியத்தில் Ae (A2) ஆகிய இரண்டு உப வகைகளைக் கொண்டிருக்கிறது.
வகை B ஆனது Bj/B1 ('j' - ஜப்பான்) மற்றும் Ba/B2 ('a' - ஆசியா) ஆகிய இரண்டு மாறுபட்ட புவியியலுக்குரிய பரவலைக் கொண்டிருக்கிறது. வகை Ba ஆனது மேலும் நான்கு உயிரினக் கிளைகளாக (B2 - B4) உட்பிரிவுகளைக் கொண்டிருக்கிறது.
வகை C ஆனது தென்-கிழக்கு ஆசியாவில் Cs (C1) மற்றும் கிழக்கு ஆசியாவில் Ce (C2) ஆகிய இரண்டு புவியியலுக்குரிய உட்பிரிவுகள் இருக்கின்றன. C உட்பிரிவுகள் ஐந்து உயிரினக் கிளைகளாகப் (C1 - C5) பிரிக்கப்படுகின்றன. ஆறாவது உயிரினக் கிளை (C6) பிலிப்பைன்ஸில் விவரிக்கப்பட்டுள்ளது. ஆனால் தற்போது ஒரேயொரு பிரித்தெடுப்பில் மட்டுமே.[19] வகை C1 வியட்நாம், மியான்மர் மற்றும் தாய்லாந்து ஆகியவற்றுடன் தொடர்புடையதாக இருக்கிறது; வகை C2 ஜப்பான், கொரியா மற்றும் சீனா ஆகியவற்றுடனும், வகை C3 நியூ கலடோனியா மற்றும் பாலினேசியா ஆகியவற்றுடனும், C4 ஆஸ்திரேலியாவுடனும் மற்றும் C5 பிலிப்பைன்ஸ் உடனும் தொடர்புடையதாக இருக்கின்றன. மேலுமொரு உட்பிரிவு பாப்புவா, இந்தோனேசியா ஆகியவற்றில் விவரிக்கப்பட்டிருக்கிறது.[20]
வகை D ஆனது 7 உட்பிரிவுகளாகப் (D1 - D7) பிரிக்கப்படுகின்றது.
வகை F ஆனது 4 உட்பிரிவுகளாகப் (F1 - F4) பிரிக்கப்படுகின்றது. F1 ஆனது மேலும் 1a மற்றும் 1b ஆகிய உட்பிரிவுகளாகப் பிரிக்கப்பட்டுள்ளது. வெனுசுலாவில் F1, F2 மற்றும் F3 ஆகிய உட்பிரிவுகள் கிழக்கு மற்றும் மேற்கு அமெரிண்டியன்ஸில் காணப்படுகின்றன. தென் அமெரிண்டியர்களுக்கு இடையில் F3 மட்டுமே கண்டறியப்பட்டது. Ia, III மற்றும் IV உட்பிரிவுகள் வரையறுக்கப்பட்ட புவியியலுக்குரிய பரவலை (முறையே மத்திய அமெரிக்கா, தென் அமெரிக்காவின் வட மற்றும் தென் பகுதிகள்) கொண்டிருக்கின்றன. அதே சமயம் Ib மற்றும் II உயிரினக் கிளைகள் முறையே வடக்கு தென் அமெரிக்கா தவிர்த்து அனைத்து அமெரிக்காக்கள் மற்றும் வட அமெரிக்காவில் காணப்படுகின்றன்.
வாழ்க்கைச் சுழற்சி
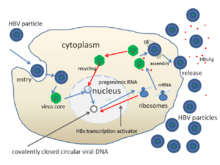
ஈரலழற்சி பி தீநுண்மத்தின் வாழ்க்கைச் சுழற்சி மிகவும் சிக்கலானது. ஈரலழற்சி பி ஆனது நேர்மாறு படியெடுத்தலை அதன் பிரதிசெய்கைச் செயல்பாட்டின் ஒரு பகுதியாகப் பயன்படுத்தும் சில அறிந்த பின்திரும்பும் நச்சுயிரி சார்ந்ததல்லாத தீநுண்மமாக இருக்கிறது.
- இணைப்பு
- இந்த தீநுண்மமானது செல்லின் புறப்பரப்பு மீது ஏற்பியில் கட்டுப்படுவதன் மூலமாக கலத்தினுள் நுழைகிறது. மேலும் என்டோசைடோசிஸ் மூலமாக அது நுழைகிறது. கல புறப்பரப்பு ஏற்பியை இன்னும் கண்டறிய வேண்டியிருக்கிறது. எனினும் இது செரைன் புரதநொதிச்சத்து வினைத்தடுப்பான்களின் ஒவல்புமின் குடும்பத்தின் உறுப்பினராக இருக்கலாம் எனக்கருதப்படுகிறது.[1]
- ஊடுருவல்
- இந்தத் தீநுண்ம மென்படலம் பின்னர் வழங்கிக் கலத்தின் மென்படலத்துடன் இணைந்து, mRNA மற்றும் அடிப்படைப் புரதங்களை குழியவுருவினுள் வெளியிடுகின்றது.
- உறை நீக்குதல்
- வழங்கி நொதி மூலமாக உருவாகும் ஆர்.என்.ஏ வழியாக தீநுண்மம் பெருக்கமடைவதன் காரணமாக, நச்சுயிரி சார்ந்த மரபுத்திகுதி சார்ந்த டி.என்.ஏவானது சாபரான்கள் என்று அழைக்கப்படும் வழங்கி புரதங்கள் மூலமாக கல அணுக்கருவுக்கு பரிமாற்றப்பட வேண்டும். பகுதியளவு இரட்டை இழையிடப்பட்ட நச்சுயிரி சார்ந்த டி.என்.ஏவில் இருந்து பிரிந்த அடிப்படைப் புரதங்கள் பின்னர் முழுமையான இரட்டை இழையிடப்பட்டதாக மாற்றப்பட்டு, நான்கு நச்சுயிரி சார்ந்த mRNAக்களின் படியெடுத்தலுக்கான வார்ப்புருவாகச் செயல்படும் சக இணைப்பில் நெருங்கிய சுழற்சி டி.என்.ஏ (cccDNA) ஆக மாற்றப்படுகின்றன.
- பிரதிசெய்கை
- மிகவும் பெரிய mRNA ஆனது (இது நச்சுயிரி சார்ந்த மரபுத்தொகுதியைக் காட்டிலும் பெரியது) மரபுத்தொகுதியின் புதிய நகல்களை உருவாக்குவதற்கு, கேப்சிட் அடிப்படைப் புரதத்தை உருவாக்குவதற்கு மற்றும் நச்சுயிரி சார்ந்த டி.என்.ஏ பாலிமரேசுவை உருவாக்குவதற்குப் பயன்படுத்தப்படுகின்றன.
- கூட்டுச்சேரல்
- இந்த நான்கு நச்சுயிரி சார்ந்த படிகளும் கூடுதல் செயல்பாட்டிற்கு உட்படுகின்றன. மேலும் கலத்திலிருந்து வெளியிடப்படும் அல்லது அணுக்கருவிற்குத் திரும்பும் சந்ததி முதிர்ந்த நச்சுயிரிகளை உருவாக்குவதைத் தொடர்கின்றன. பின்னர் மேலும் அதிகமான நகல்களை உருவாக்குவதற்காக மறுசுழற்சியைத் தொடர்கின்றன.[13][21]
- வெளியீடு
- நீண்ட mRNA பின்னர் அதன் நேர்மாறு படியெடுத்தல் நடவடிக்கை வழியாக முதிர்ந்த நச்சுயிரி P புரதமானது டி.என்.ஏ ஐத் தொகுக்கின்ற இடமான குழியவுருவுக்கு அனுப்பப்படுகிறது.
குறிப்புதவிகள்
- Hunt, Richard (2007-11-21). "Hepatitis viruses". University of Southern California, Department of Pathology and Microbiology. பார்த்த நாள் 2008-03-13.
- Hassan MM, Li D, El-Deeb AS, et al. (October 2008). "Association between hepatitis B virus and pancreatic cancer". J. Clin. Oncol. 26 (28): 4557–62. doi:10.1200/JCO.2008.17.3526. பப்மெட்:18824707. பப்மெட் சென்ட்ரல்:2562875. http://www.jco.org/cgi/pmidlookup?view=long&pmid=18824707.
- Schwalbe M, Ohlenschläger O, Marchanka A, et al. (March 2008). "Solution structure of stem-loop alpha of the hepatitis B virus post-transcriptional regulatory element". Nucleic Acids Res. 36 (5): 1681–9. doi:10.1093/nar/gkn006. பப்மெட்:18263618. பப்மெட் சென்ட்ரல்:2275152. http://nar.oxfordjournals.org/cgi/pmidlookup?view=long&pmid=18263618.
- Mason, W.S. et al. (2008-07-08). "00.030. Hepadnaviridae - ICTVdB Index of Viruses". International Committee on Taxonomy of Viruses. பார்த்த நாள் 2009-03-13.
- Kramvis A, Kew M, François G (2005). "Hepatitis B virus genotypes". Vaccine 23 (19): 2409–23. doi:10.1016/j.vaccine.2004.10.045. பப்மெட்:15752827.
- Magnius LO, Norder H (1995). "Subtypes, genotypes and molecular epidemiology of the hepatitis B virus as reflected by sequence variability of the S-gene". Intervirology 38 (1-2): 24–34. பப்மெட்:8666521.
- Zuckerman AJ (1996). Hepatitis Viruses. In: Baron's Medical Microbiology (Baron S et al., eds.) (4th ). Univ of Texas Medical Branch. பன்னாட்டுத் தரப்புத்தக எண்:0-9631172-1-1. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mmed.section.3738.
- Locarnini S (2004). "Molecular virology of hepatitis B virus". Semin. Liver Dis. 24 Suppl 1: 3–10. doi:10.1055/s-2004-828672. பப்மெட்:15192795.
- Howard CR (1986). "The biology of hepadnaviruses". J. Gen. Virol. 67 ( Pt 7): 1215–35. doi:10.1099/0022-1317-67-7-1215. பப்மெட்:3014045.
- Guo GH, Tan DM, Zhu PA, Liu F (February 2009). "Hepatitis B virus X protein promotes proliferation and upregulates TGF-beta1 and CTGF in human hepatic stellate cell line, LX-2". Hbpd Int 8 (1): 59–64. பப்மெட்:19208517. http://www.hbpdint.com/text.asp?id=1196.
- Chai N, Chang HE, Nicolas E, Han Z, Jarnik M, Taylor J (August 2008). "Properties of subviral particles of hepatitis B virus". J. Virol. 82 (16): 7812–7. doi:10.1128/JVI.00561-08. பப்மெட்:18524834. பப்மெட் சென்ட்ரல்:2519590. http://jvi.asm.org/cgi/pmidlookup?view=long&pmid=18524834.
- Kay A, Zoulim F (2007). "Hepatitis B virus genetic variability and evolution". Virus Res. 127 (2): 164–76. doi:10.1016/j.virusres.2007.02.021. பப்மெட்:17383765.
- Beck J, Nassal M (2007). "Hepatitis B virus replication". World J. Gastroenterol. 13 (1): 48–64. பப்மெட்:17206754.
- Bouchard MJ, Schneider RJ (2004). "The enigmatic X gene of hepatitis B virus". J. Virol. 78 (23): 12725–34. doi:10.1128/JVI.78.23.12725-12734.2004. பப்மெட்:15542625.
- Olinger CM, Jutavijittum P, Hübschen JM, et al. (November 2008). "Possible new hepatitis B virus genotype, southeast Asia". Emerging Infect. Dis. 14 (11): 1777–80. doi:10.3201/eid1411.080437. பப்மெட்:18976569. பப்மெட் சென்ட்ரல்:2630741. http://www.cdc.gov/eid/content/14/11/1777.htm.
- Kurbanov F, Tanaka Y, Kramvis A, Simmonds P, Mizokami M (August 2008). "When should "I" consider a new hepatitis B virus genotype?". J. Virol. 82 (16): 8241–2. doi:10.1128/JVI.00793-08. பப்மெட்:18663008. பப்மெட் சென்ட்ரல்:2519592. http://jvi.asm.org/cgi/pmidlookup?view=long&pmid=18663008.
- Palumbo E (2007). "Hepatitis B genotypes and response to antiviral therapy: a review". Am J Ther 14 (3): 306–9. doi:10.1097/01.pap.0000249927.67907.eb. பப்மெட்:17515708. http://meta.wkhealth.com/pt/pt-core/template-journal/lwwgateway/media/landingpage.htm?an=00045391-200705000-00016.
- Mahtab MA, Rahman S, Khan M, Karim F (October 2008). "Hepatitis B virus genotypes: an overview". Hbpd Int 7 (5): 457–64. பப்மெட்:18842489. http://www.hbpdint.com/text.asp?id=1142.
- Cavinta L., Sun J., May A., Yin J., von Meltzer M., Radtke M., Barzaga N.G., Cao G., Schaefer S.(2009) A new isolate of hepatitis B virus from the Philippines possibly representing a new subgenotype C6. J. Med. Virol. 81(6):983-987
- Lusida M.I., Nugrahaputra V.E., Soetjipto, Handajani R., Nagano-Fujii M., Sasayama M., Utsumi T., Hotta H.(2008) Novel subgenotypes of hepatitis B virus genotypes C and D in Papua, Indonesia. J. Clin. Microbiol. 46(7):2160-2166
- Bruss V (2007). "Hepatitis B virus morphogenesis". World J. Gastroenterol. 13 (1): 65–73. பப்மெட்:17206755.