Liver sinusoidal endothelial cell
Liver sinusoidal endothelial cells (LSECs) form the lining of the smallest blood vessels in the liver, also called the hepatic sinusoids. LSECs are highly specialized endothelial cells with characteristic morphology and function. They constitute an important part of the reticuloendothelial system (RES).
| Liver sinusoidal endothelial cell | |
|---|---|
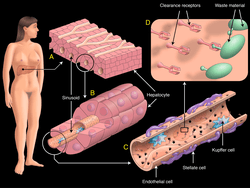 Schematic view of the cells in liver sinusoids. A. Liver segment with numerous sinusoids, surrounded by hepatocytes. B. Liver sinusoidal endothelial cells (LSECs) lining the wall of the liver sinusoid. C. Kupffer cells (blue) distributed along the luminal aspect of the sinusoid, and hepatic stellate cells (violet) located along the abluminal aspect. D. Surface of an LSEC with numerous receptors for clearance of circulating waste (green). | |
| Details | |
| System | Circulatory system |
| Location | blood vessels of liver |
| Anatomical terms of microanatomy | |
Structure
Although the LSECs make up only about 3% of the total liver cell volume, their surface in a normal adult human liver is about 210 m2, or nearly the size of a tennis court (calculated from data published in [1]. The LSEC structure differs from other endothelia. 1. The cells contain many open pores, or fenestrae, with diameters from 100 to 150 nm. The fenestrae occupy 20% of the LSEC surface and are arranged in groups referred to as “sieve plates” [2]. Filtering fluid between the sinusoidal lumen and the space of Disse the fenestrae are crucial for lipoprotein traffic between the hepatocytes and the sinusoidal lumen [3]. 2. The LSECs lack an organized basal lamina [4]. 3. The LSECs contain 45% and 17% of the liver’s total mass of pinocytic vesicles and lysosomes, resp. (1), and contain twice as many clathrin-coated pits per membrane unit, compared with two other major liver cells, Kupffer cells and hepatocytes[5], reflecting the high capacity clathrin-mediated endocytic activity of LSECs.
Physiological functions
LSECs play a central role in the clearance of blood borne waste. The cells express endocytosis receptors that mediate extremely rapid internalization of waste molecules. In rat it has been shown that LSECs express scavenger receptors (SR) class A, B, E and H [6]. The latter exists as stabilin-1 (SR-H1) and stabilin-2 (SR-H2) in LSECs. In the liver stabilin-2, the most important SR on LSECs, is uniquely expressed in these cells. Moreover, LSECs also express high levels of the macrophage mannose receptor (MMR) [7] and the Fc-gamma receptor IIb2 (FcRIIb2) [8], both highly active in clathrin-mediated endocytosis just like the two stabilins. Other important receptors on LSECs are L-SIGN (liver/lymph node-specific ICAM-3 grabbing nonintegrin) [9], LSECtin (liver and lymph node sinusoidal endothelial cell C-type lectin) [10], Lyve-1 (lymphatic vessel endothelial hyaluronan receptor‐1) [11], and LRP‐1 (low‐density lipoprotein receptor‐related protein‐1) [12].
The capacity of LSECs as scavengers of blood borne waste assigns an important role of these cells in innate immunity. The abundant expression of receptors such as the endocytic FcRIIb2 and pattern recognition receptors (PRRs) i.e. toll like receptors (TLRs), MMR and SRs, as well as the high expression of inflammasome molecules NLRP-1, NLRP-3, and AIM2 [13] point to innate immune functions of LSECs. In addition, LSECs display features of adaptive immunity, contributing to hepatic immune tolerance [14].
Pathobiology
Liver fibrosis
LSECs have been reported to play a role in the development of liver fibrosis. Liver fibrosis is associated with decreased LSEC fenestration, and appearance of an organized basal lamina in the space of Disse, a process called capillarization, which precedes the onset of liver fibrosis [15]. Normally differentiated LSECs prevent hepatic stellate cell activation and promote reversion to quiescence, whereas capillarized LSECs do not [16].
Atherosclerosis
Chylomicrons produced by the intestinal epithelial cells from dietary lipids have diameter up to 1000 nm which prevents them from passing through the LSEC fenestrae [17]. The size of circulating chylomicrons is gradually reduced to chylomicron remnants by lipoprotein lipase on endothelial cells of systemic capillaries. When the chylomicron remnants become small enough (30‐80 nm), they pass through the LSEC fenestrations, leading to their metabolism in hepatocytes. Reduced porosity, as in liver cirrhosis, diabetes mellitus or old age may lead to prolonged postprandial lipoproteinemia and increased circulatory cholesterol levels, with increased risk for development of atherosclerosis [18].
Autoimmunity
It has been suggested that reduced Fc receptor function in humans, causing increased circulating levels of soluble immune complexes is important in the etiology of autoimmune diseases such as systemic lupus erythematosus (SLE) and Sjögren’s syndrome [19]. Moreover, the observation that small soluble IgG-antigen immune complexes are cleared in the mouse mainly via the LSEC FcRIIb2 (8), along with the observation that deletion of same receptor causes spontaneous auto-immunity and SLE-like disease in mice [20], point to a pivotal role of LSEC FcRIIb2 in the disease mechanism of SLE. Furthermore, the finding that scavenging of blood borne DNAs is chiefly by SR-mediated uptake in LSECs [21], along with the fact that SLE is associated with generation of anti-DNA antibody, lend additional support to the hypothesis that LSECs participate in the onset of SLE.
Liver toxicology
LSECs may sometimes be the initial target of injury in a condition referred to as sinusoidal obstruction syndrome (SOS, formerly hepatic veno-occlusive disease, VOD), which is described as a change of the sinusoid that may lead to hepatocyte hypoxia, with liver dysfunction and disruption of the portal circulation. Major causes of SOS are dietary ingestion of pyrrolizidine alkaloids, treatment with several chemotherapeutic drugs, and acetaminophen [22]. Moreover, since LSECs are geared to (generally unwanted) active blood clearance of large molecule compounds and nano formulations (7) these cells may be easily intoxicated by off-target mechanisms, causing subsequent hepatotoxicity [23].
Origin and renewal of LSECs
Normal LSEC turnover is maintained by the liver resident population of LSEC progenitor cells; in addition, recruitment of bone marrow derived cells contributes to replenish the LSEC population when needed [24].
History
By the end of the 19th and beginning of the 20th centuries researchers observed avid accumulation of intravenously administered colloidal vital dyes in cells lining the sinusoids of some tissues, with the highest uptake in the littoral cells of the hepatic sinusoids. These very active blood clearance cells were collectively named “The reticuloendothelial system”, or RES [25]. For several decades it was believed that the cells comprising the RES were the macrophages of the mononuclear phagocyte system (MPS). Hence the hepatic clearance of circulating waste was attributed to the liver macrophages, or Kupffer cells [26]. However, by a recent re-investigation of the original vital stain experiments carried out 100-140 years ago [27] it was concluded that the vital stain accumulated mainly in LSECs. It is increasingly accepted that LSECs and Kupffer cells play complementary roles in the hepatic blood clearance process, referred to as the dual cell principle of waste clearance (6): LSECs clear macromolecules and nanoparticles roughly <200 nm by clathrin-mediated endocytosis whereas Kupffer cells clear larger particles >200 nm by phagocytosis.
LSECs in a comparative context
All vertebrates carry a population of endothelial cells that are remarkably active in blood clearance of macromolecules and nano-substances. The great majority of these cells are located in the liver sinusoids of land-based vertebrates (mammals, birds, reptiles and amphibia). However, in bony fishes, these specialized endothelial cells are located either in the heart endocardium or in endothelial cells of the kidney sinusoidal lining, depending on the fish species. In cartilaginous fishes and the jawless fishes, these endothelial cells constitute the lining of the gill capillaries. The name scavenger endothelial cells (SECs) has been coined to denote the endothelial cells in vertebrates that are geared to blood clearance [28]. Thus, LSECs in mammals and other land-based vertebrates are a member of the vertebrate SEC family.
References
- Blouin, A; Bolender, RP; Weibel, ER (February 1977). "Distribution of organelles and membranes between hepatocytes and nonhepatocytes in the rat liver parenchyma. A stereological study". The Journal of Cell Biology. 72 (2): 441–55. doi:10.1083/jcb.72.2.441. PMC 2110997. PMID 833203.
- Cogger, Victoria C.; Couteur, David G. Le (2009). "Fenestrations in the Liver Sinusoidal Endothelial Cell". The Liver. John Wiley & Sons, Ltd. pp. 389–406. doi:10.1002/9780470747919.ch27. ISBN 9780470747919.
- Fraser, R; Cogger, VC; Dobbs, B; Jamieson, H; Warren, A; Hilmer, SN; Le Couteur, DG (April 2012). "The liver sieve and atherosclerosis". Pathology. 44 (3): 181–6. doi:10.1097/PAT.0b013e328351bcc8. PMID 22406487.
- Wisse, E (March 1972). "An ultrastructural characterization of the endothelial cell in the rat liver sinusoid under normal and various experimental conditions, as a contribution to the distinction between endothelial and Kupffer cells". Journal of Ultrastructure Research. 38 (5): 528–62. doi:10.1016/0022-5320(72)90089-5. PMID 4335119.
- Kjeken, R; Mousavi, SA; Brech, A; Gjøen, T; Berg, T (May 2001). "Fluid phase endocytosis of [125I]iodixanol in rat liver parenchymal, endothelial and Kupffer cells". Cell and Tissue Research. 304 (2): 221–30. doi:10.1007/s004410100348. PMID 11396716.
- Sørensen, KK; McCourt, P; Berg, T; Crossley, C; Le Couteur, D; Wake, K; Smedsrød, B (15 December 2012). "The scavenger endothelial cell: a new player in homeostasis and immunity". American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 303 (12): R1217–30. doi:10.1152/ajpregu.00686.2011. PMID 23076875.
- Sørensen, KK; Simon-Santamaria, J; McCuskey, RS; Smedsrød, B (20 September 2015). "Liver Sinusoidal Endothelial Cells". Comprehensive Physiology. 5 (4): 1751–74. doi:10.1002/cphy.c140078. PMID 26426467.
- Mousavi, SA; Sporstøl, M; Fladeby, C; Kjeken, R; Barois, N; Berg, T (September 2007). "Receptor-mediated endocytosis of immune complexes in rat liver sinusoidal endothelial cells is mediated by FcgammaRIIb2". Hepatology. 46 (3): 871–84. doi:10.1002/hep.21748. PMID 17680646.
- Pöhlmann, S; Soilleux, EJ; Baribaud, F; Leslie, GJ; Morris, LS; Trowsdale, J; Lee, B; Coleman, N; Doms, RW (27 February 2001). "DC-SIGNR, a DC-SIGN homologue expressed in endothelial cells, binds to human and simian immunodeficiency viruses and activates infection in trans". Proceedings of the National Academy of Sciences of the United States of America. 98 (5): 2670–5. Bibcode:2001PNAS...98.2670P. doi:10.1073/pnas.051631398. PMC 30196. PMID 11226297.
- Liu, W; Tang, L; Zhang, G; Wei, H; Cui, Y; Guo, L; Gou, Z; Chen, X; Jiang, D; Zhu, Y; Kang, G; He, F (30 April 2004). "Characterization of a novel C-type lectin-like gene, LSECtin: demonstration of carbohydrate binding and expression in sinusoidal endothelial cells of liver and lymph node". The Journal of Biological Chemistry. 279 (18): 18748–58. doi:10.1074/jbc.M311227200. PMID 14711836.
- Martens, JH; Kzhyshkowska, J; Falkowski-Hansen, M; Schledzewski, K; Gratchev, A; Mansmann, U; Schmuttermaier, C; Dippel, E; Koenen, W; Riedel, F; Sankala, M; Tryggvason, K; Kobzik, L; Moldenhauer, G; Arnold, B; Goerdt, S (March 2006). "Differential expression of a gene signature for scavenger/lectin receptors by endothelial cells and macrophages in human lymph node sinuses, the primary sites of regional metastasis". The Journal of Pathology. 208 (4): 574–89. doi:10.1002/path.1921. PMID 16440291.
- Øie, CI; Appa, RS; Hilden, I; Petersen, HH; Gruhler, A; Smedsrød, B; Hansen, JB (December 2011). "Rat liver sinusoidal endothelial cells (LSECs) express functional low density lipoprotein receptor-related protein-1 (LRP-1)". Journal of Hepatology. 55 (6): 1346–52. doi:10.1016/j.jhep.2011.03.013. hdl:10037/4078. PMID 21703209.
- Boaru, SG; Borkham-Kamphorst, E; Tihaa, L; Haas, U; Weiskirchen, R (28 November 2012). "Expression analysis of inflammasomes in experimental models of inflammatory and fibrotic liver disease". Journal of Inflammation (London, England). 9 (1): 49. doi:10.1186/1476-9255-9-49. PMC 3599703. PMID 23192004.
- Knolle, PA; Wohlleber, D (May 2016). "Immunological functions of liver sinusoidal endothelial cells". Cellular & Molecular Immunology. 13 (3): 347–53. doi:10.1038/cmi.2016.5. PMC 4856811. PMID 27041636.
- DeLeve, LD (May 2015). "Liver sinusoidal endothelial cells in hepatic fibrosis". Hepatology. 61 (5): 1740–6. doi:10.1002/hep.27376. PMC 4333127. PMID 25131509.
- Xie, G; Wang, X; Wang, L; Wang, L; Atkinson, RD; Kanel, GC; Gaarde, WA; Deleve, LD (April 2012). "Role of differentiation of liver sinusoidal endothelial cells in progression and regression of hepatic fibrosis in rats". Gastroenterology. 142 (4): 918–927.e6. doi:10.1053/j.gastro.2011.12.017. PMC 3618963. PMID 22178212.
- Naito, M; Wisse, E (10 July 1978). "Filtration effect of endothelial fenestrations on chylomicron transport in neonatal rat liver sinusoids". Cell and Tissue Research. 190 (3): 371–82. doi:10.1007/bf00219553. PMID 567529.
- Fraser, R; Cogger, VC; Dobbs, B; Jamieson, H; Warren, A; Hilmer, SN; Le Couteur, DG (April 2012). "The liver sieve and atherosclerosis". Pathology. 44 (3): 181–6. doi:10.1097/PAT.0b013e328351bcc8. PMID 22406487.
- Frank, MM; Lawley, TJ; Hamburger, MI; Brown, EJ (February 1983). "NIH Conference: Immunoglobulin G Fc receptor-mediated clearance in autoimmune diseases". Annals of Internal Medicine. 98 (2): 206–18. doi:10.7326/0003-4819-98-2-218. PMID 6824256.
- Ahmed, SS; Muro, H; Nishimura, M; Kosugi, I; Tsutsi, Y; Shirasawa, H (July 1995). "Fc receptors in liver sinusoidal endothelial cells in NZB/W F1 lupus mice: a histological analysis using soluble immunoglobulin G-immune complexes and a monoclonal antibody (2.4G2)". Hepatology. 22 (1): 316–24. doi:10.1002/hep.1840220143. hdl:10271/1047. PMID 7541388.
- Hisazumi, J; Kobayashi, N; Nishikawa, M; Takakura, Y (July 2004). "Significant role of liver sinusoidal endothelial cells in hepatic uptake and degradation of naked plasmid DNA after intravenous injection". Pharmaceutical Research. 21 (7): 1223–8. doi:10.1023/B:PHAM.0000033009.17594.e5. PMID 15290863.
- DeLeve, LD (November 2007). "Hepatic microvasculature in liver injury". Seminars in Liver Disease. 27 (4): 390–400. doi:10.1055/s-2007-991515. PMID 17979075.
- Godfrey, C; Desviat, LR; Smedsrød, B; Piétri-Rouxel, F; Denti, MA; Disterer, P; Lorain, S; Nogales-Gadea, G; Sardone, V; Anwar, R; El Andaloussi, S; Lehto, T; Khoo, B; Brolin, C; van Roon-Mom, WM; Goyenvalle, A; Aartsma-Rus, A; Arechavala-Gomeza, V (May 2017). "Delivery is key: lessons learnt from developing splice-switching antisense therapies". EMBO Molecular Medicine. 9 (5): 545–557. doi:10.15252/emmm.201607199. PMC 5412803. PMID 28289078.
- DeLeve, LD (May 2013). "Liver sinusoidal endothelial cells and liver regeneration". The Journal of Clinical Investigation. 123 (5): 1861–6. doi:10.1172/JCI66025. PMC 3635729. PMID 23635783.
- Aschoff, L. (1924). "Das reticulo-endotheliale System". Ergebnisse der Inneren Medizin und Kinderheilkunde: Sechsundzwanzigster Band (in German). Springer Berlin Heidelberg: 1–118. doi:10.1007/978-3-642-90639-8_1. ISBN 978-3-642-88784-0.
- van Furth, R; Cohn, ZA; Hirsch, JG; Humphrey, JH; Spector, WG; Langevoort, HL (1972). "The mononuclear phagocyte system: a new classification of macrophages, monocytes, and their precursor cells". Bulletin of the World Health Organization. 46 (6): 845–52. PMC 2480884. PMID 4538544.
- Wake, K; Kawai, Y; Smedsrød, B (2001). "Re-evaluation of the reticulo-endothelial system". Italian Journal of Anatomy and Embryology. 106 (2 Suppl 1): 261–9. PMID 11729964.
- Seternes, T; Sørensen, K; Smedsrød, B (28 May 2002). "Scavenger endothelial cells of vertebrates: a nonperipheral leukocyte system for high-capacity elimination of waste macromolecules". Proceedings of the National Academy of Sciences of the United States of America. 99 (11): 7594–7. Bibcode:2002PNAS...99.7594S. doi:10.1073/pnas.102173299. PMC 124295. PMID 12032328.
External links
- Liver Research at VBRG Liver Sinusoidal Endothelial Cell Biology