Neisseria
Neisseria is a large genus of bacteria that colonize the mucosal surfaces of many animals. Of the 11 species that colonize humans, only two are pathogens, N. meningitidis and N. gonorrhoeae. Most gonococcal infections are asymptomatic and self-resolving, and epidemic strains of the meningococcus may be carried in >95% of a population where systemic disease occurs at <1% prevalence.
| Neisseria | |
|---|---|
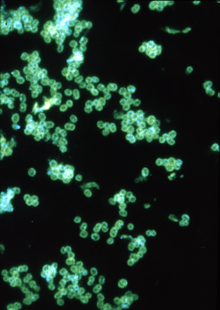 | |
| Fluorescent antibody stain of Neisseria gonorrhoeae. | |
| Scientific classification | |
| Domain: | Bacteria |
| Phylum: | Proteobacteria |
| Class: | Betaproteobacteria |
| Order: | Neisseriales |
| Family: | Neisseriaceae |
| Genus: | Neisseria Trevisan, 1885 |
| Species | |
|
N. animalis | |
Neisseria species are Gram-negative bacteria included among the proteobacteria, a large group of Gram-negative forms. Neisseria diplococci resemble coffee beans when viewed microscopically.[1]
History
The genus Neisseria is named after the German bacteriologist Albert Neisser, who in 1879 discovered its first example, Neisseria gonorrhoeae, the pathogen which causes the human disease gonorrhea. Neisser also co-discovered the pathogen that causes leprosy, Mycobacterium leprae. These discoveries were made possible by the development of new staining techniques which he helped to develop.
Classification
Pathogens
Species of this genus (family Neisseriaceae) of parasitic bacteria grow in pairs and occasionally tetrads, and thrive best at 98.6 °F (37 °C) in the animal body or serum media.
The genus includes:
- N. gonorrhoeae (also called the gonococcus) causes gonorrhea.
- N. meningitidis (also called the meningococcus) is one of the most common causes of bacterial meningitis and the causative agent of meningococcal septicaemia.
These two species have the ability of 'breaching' the barrier. Local cytokines of the area become secreted to initiate an immune response. However, neutrophils are not able to do their job due to the ability of Neisseria to invade and replicate within neutrophils, as well avoiding phagocytosis and being killed by complement by resisting opsonization by antibodies, which target the pathogen for destruction. Neisseria species are also able to alter their antigens to avoid being engulfed by a process called antigenic variation, which is observed primarily in surface-located molecules. The pathogenic species along with some commensal species, have type IV pili which serve multiple functions for this organism. Some functions of the type IV pili include: mediating attachment to various cells and tissues, twitching motility, natural competence, microcolony formation, extensive intrastrain phase, and antigenic variation.
Neisseria bacteria have also been shown to be an important factor in the early stages of canine plaque development.[2]
Nonpathogens
This genus also contains several, believed to be commensal, or nonpathogenic, species:
- Neisseria bacilliformis
- Neisseria cinerea
- Neisseria elongata
- Neisseria flavescens
- Neisseria lactamica
- Neisseria macacae
- Neisseria mucosa
- Neisseria oralis
- Neisseria polysaccharea
- Neisseria sicca
- Neisseria subflava
- Neisseria flava
However, some of these can be associated with disease.[4][5]
Biochemical identification
All the medically significant species of Neisseria are positive for both catalase and oxidase. Different Neisseria species can be identified by the sets of sugars from which they will produce acid. For example, N. gonorrhoeae makes acid from only glucose, but N. meningitidis produces acid from both glucose and maltose.
Polysaccharide capsule. N. meningitidis has a polysaccharide capsule that surrounds the outer membrane of the bacterium and protects against soluble immune effector mechanisms within the serum. It is considered to be an essential virulence factor for the bacteria.[6] N. gonorrhoeae possesses no such capsule. Instead of having the usual lipopolysaccharide (LPS), this bacterium, whether a pathogenic or commensal species, has a lipooligosaccharide (LOS) which consists of a core polysaccharide and lipid A. It functions as an endotoxin, protects against antimicrobial peptides, and adheres to the asialoglycoprotein receptor on urethral cells. LOS is highly stimulatory to the human immune system. LOS sialylation (by the enzyme's Lst) prevents complement deposition and phagocytosis by neutrophils. LOS modification by phosphoethanolamine (by the enzyme LptA) provides resistance to antimicrobial peptides and complement. Strains of the same species have the ability to produce different LOS glycoforms.
Genomes
The genomes of at least 10 Neisseria species have been completely sequenced.[3] The best-studied species are N. meningitidis with more than 70 strains and N. gonorrhoeae with at least 10 strains completely sequenced. Other complete genomes are available for N. elongata, N. lactamica,[7] and N. weaveri. Whole genome shotgun sequences are available for hundreds of other species and strains.[8] N. meningitidis encodes 2,440 to 2,854 proteins while N. gonorrhoeae encodes from 2,603 to 2,871 proteins. N. weaveri (strain NCTC 13585) has the smallest known genome with only 2,060 encoded proteins[9] although N. meningitidis MC58 has been reported to have only 2049 genes.[3] The genomes are generally quite similar. For example, when the genome of N. gonorrhoeae (strain FA1090) is compared to that of N. meningitidis (strain H44/76) 68% of their genes are shared.[8]
| Genome properties of Neisseria sp.[3] | ||
|---|---|---|
| species | Size (bp) | gene number |
| N. elongata | 2,260,105 | 2589 |
| N. sicca | 2,786,309 | 2842 |
| N. mucosa | 2,542,952 | 2594 |
| N. subflava | 2,288,219 | 2303 |
| N. flavescens | 2,199,447 | 2240 |
| N. cinerea | 1,876,338 | 2050 |
| N. polysaccharea | 2,043,594 | 2268 |
| N. lactamica 23970 | 2,148,211 | 2359 |
| N. gonorrhoeae FA1090 | 2,153,922 | 2002 |
| N. meningitidis MC58 | 2,184,406 | 2049 |
Iron acquisition
Iron is absolutely required by all life forms, playing a critical role in a number of essential processes. Free iron, at least what would be readily available to a microbial pathogen, practically does not exist in animals. In vertebrates, the majority of iron is stored inside cells in complex with either ferritin or hemoglobin. Extracellular iron is found in body fluids complexed to either transferrin or lactoferrin.
Pathogens acquire iron by two different strategies
- ‘Siderophore’-mediated iron uptake involves outcompeting transferrin and/or lactoferrin for iron binding. Iron-bound siderophores are then taken into the bacterium by specific receptors.
- Direct uptake of iron-bound host proteins involves the bacteria possessing a high affinity for transferrin, lactoferrin, and hemoglobin (the approach used by the pathogenic Neiserria spp.).
Receptors: HmbRm, HpuA, and HpuB are receptors for haptoglobin-haemoglobin. LbpAB is a receptor for human lactoferrin. TbpAB (Tbp1-Tbp2) is a receptor for human transferrin. All of these receptors are used for iron acquisition for both pathogenic and commensal species.
Vaccine
Diseases caused by N. meningitidis and N. gonorrhoeae are significant health problems worldwide, the control of which is largely dependent on the availability and widespread use of comprehensive meningococcal and gonococcal vaccines. Development of neisserial vaccines has been challenging due to the nature of these organisms, in particular the heterogeneity, variability and/or poor immunogenicity of their outer surface components. As strictly human pathogens, they are highly adapted to the host environment, but have evolved several mechanisms to remain adaptable to changing microenvironments and avoid elimination by the host immune system. Currently, serogroup A, B, C, Y, and W-135 meningococcal infections can be prevented by vaccines.[10] However, the prospect of developing a gonococcal vaccine is remote.[11]
Antibiotic resistance
The acquisition of cephalosporin resistance in N. gonorrhoeae, particularly ceftriaxone resistance, has greatly complicated the treatment of gonorrhea, with the gonococcus now being classified as a "superbug".[12]
Genetic transformation
Genetic transformation is the process by which a recipient bacterial cell takes up DNA from a neighboring cell and integrates this DNA into the recipient’s genome by recombination. In N. meningitidis and N. gonorrhoeae, DNA transformation requires the presence of short DNA sequences (9-10 monomers residing in coding regions) of the donor DNA. These sequences are called DNA uptake sequences (DUSs). Specific recognition of DUSs is mediated by a type IV pilin.[13] Davidsen et al.[14] reported that in N. meningitidis and N. gonorrhoeae, DUSs occur at a significantly higher density in genes involved in DNA repair and recombination (as well as in restriction-modification and replication) than in other annotated gene groups. These authors proposed that the over-representation of DUS in DNA repair and recombination genes may reflect the benefit of maintaining the integrity of the DNA repair and recombination machinery by preferentially taking up genome maintenance genes that could replace their damaged counterparts in the recipient cell. Caugant and Maiden noted that the distribution of DUS is consistent with recombination being primarily a mechanism for genome repair that can occasionally result in generation of diversity, which even more occasionally, is adaptive.[15] It was also suggested by Michod et al.[16] that an important benefit of transformation in N. gonorrhoeae is recombinational repair of oxidative DNA damages caused by oxidative attack by the host’s phagocytic cells.
International Pathogenic Neisseria Conference
The International Pathogenic Neisseria Conference (IPNC), occurring every two years, is a forum for the presentation of cutting-edge research on all aspects of the genus Neisseria. This includes immunology, vaccinology, and physiology and metabolism of N. meningitidis, N. gonorrhoeae and the commensal species. The first IPNC took place in 1978, and the most recent one was in September 2016. Normally, the location of the conference switches between North America and Europe, but it took place in Australia for the first time in 2006, where the venue was located in Cairns.
References
- Ryan KJ; Ray CG, eds. (2004). Sherris Medical Microbiology (4th ed.). McGraw Hill. ISBN 978-0-8385-8529-0.
- Early Canine Plaque Biofilms: Characterization of Key Bacterial Interactions Involved in Initial Colonization of Enamel. Lucy J. Holcombe, Niran Patel, Alison Colyer, Oliver Deusch, Ciaran O’Flynn, Stephen Harris. PLOS One, 2014.
- Marri, Pradeep Reddy; Paniscus, Mary; Weyand, Nathan J.; Rendón, María A.; Calton, Christine M.; Hernández, Diana R.; Higashi, Dustin L.; Sodergren, Erica; Weinstock, George M. (2010-07-28). "Genome Sequencing Reveals Widespread Virulence Gene Exchange among Human Neisseria Species". PLOS One. 5 (7): e11835. doi:10.1371/journal.pone.0011835. ISSN 1932-6203. PMC 2911385. PMID 20676376.
- Tronel H, Chaudemanche H, Pechier N, Doutrelant L, Hoen B (May 2001). "Endocarditis due to Neisseria mucosa after tongue piercing". Clin. Microbiol. Infect. 7 (5): 275–6. doi:10.1046/j.1469-0691.2001.00241.x. PMID 11422256.
- Wolfgang, WJ; Passaretti, TV; Jose, R; Cole, J; Coorevits, A; Carpenter, AN; Jose, S; Van Landschoot, A; Izard, J; Kohlerschmidt, DJ; Vandamme, P; Dewhirst, FE; Fisher, MA; Musser, KA (April 2013). "Neisseria oralis sp. nov., isolated from healthy gingival plaque and clinical samples". International Journal of Systematic and Evolutionary Microbiology. 63 (Pt 4): 1323–8. doi:10.1099/ijs.0.041731-0. PMC 3709538. PMID 22798652.
- Ullrich, M, ed. (2009). Bacterial Polysaccharides: Current Innovations and Future Trends. Caister Academic Press. ISBN 978-1-904455-45-5.
- Minogue, T. D.; Daligault, H. A.; Davenport, K. W.; Bishop-Lilly, K. A.; Bruce, D. C.; Chain, P. S.; Chertkov, O.; Coyne, S. R.; Freitas, T. (2014-09-25). "Draft Genome Assembly of Neisseria lactamica Type Strain A7515". Genome Announcements. 2 (5): e00951–14. doi:10.1128/genomeA.00951-14. PMC 4175205. PMID 25291770.
- "Neisseria in the PATRIC database". PATRIC. 2017-02-26. Retrieved 2017-02-26.
- Alexander, Sarah; Fazal, Mohammed-Abbas; Burnett, Edward; Deheer-Graham, Ana; Oliver, Karen; Holroyd, Nancy; Parkhill, Julian; Russell, Julie E. (2016-08-25). "Complete Genome Sequence of Neisseria weaveri Strain NCTC13585". Genome Announcements. 4 (4): e00815–16. doi:10.1128/genomeA.00815-16. PMC 5000823. PMID 27563039.
- "meningococcal group B vaccine". Medscape. WebMD. Retrieved December 16, 2015.
- Seib KL, Rappuoli R (2010). "Difficulty in Developing a Neisserial Vaccine". Neisseria: Molecular Mechanisms of Pathogenesis. Caister Academic Press. ISBN 978-1-904455-51-6.
- Unemo M, Nicholas RA (December 2012). "Emergence of multidrug-resistant, extensively drug-resistant and untreatable gonorrhea". Future Microbiol. 7 (12): 1401–1422. doi:10.2217/fmb.12.117. PMC 3629839. PMID 23231489.
- Cehovin A, Simpson PJ, McDowell MA, Brown DR, Noschese R, Pallett M, Brady J, Baldwin GS, Lea SM, Matthews SJ, Pelicic V (2013). "Specific DNA recognition mediated by a type IV pilin". Proc. Natl. Acad. Sci. U.S.A. 110 (8): 3065–70. doi:10.1073/pnas.1218832110. PMC 3581936. PMID 23386723.
- Davidsen T, Rødland EA, Lagesen K, Seeberg E, Rognes T, Tønjum T (2004). "Biased distribution of DNA uptake sequences towards genome maintenance genes". Nucleic Acids Res. 32 (3): 1050–8. doi:10.1093/nar/gkh255. PMC 373393. PMID 14960717.
- Caugant DA, Maiden MC (2009). "Meningococcal carriage and disease--population biology and evolution". Vaccine. 27 (Suppl 2): B64–70. doi:10.1016/j.vaccine.2009.04.061. PMC 2719693. PMID 19464092.
- Michod RE, Bernstein H, Nedelcu AM (2008). "Adaptive value of sex in microbial pathogens". Infect. Genet. Evol. 8 (3): 267–85. doi:10.1016/j.meegid.2008.01.002. PMID 18295550.