বর্ণের দৃশ্যমানতা
'''বর্ণের দৃশ্যমানতা''' হল কোন জীব অথবা কোন মেশিনের বস্তুকে সেটি হতে প্রতিফলিত, প্রতিসরিত অথবা নিঃসরিত আলোর তরঙ্গ দৈর্ঘ্য (অথবা কম্পাঙ্ক) এর উপর ভিক্তি করে চিনতে পারা। বর্ণকে বিভিন্ন উপায়ে পরিমাপ করা যায়; প্রকৃতপক্ষে একটি মানুষের বর্ণকে উপলব্ধি করা একটা সাব্জেক্টিভ ধারা যেটা আলোর মানুষের চোখের কোন সেলের মধ্যবর্তী বিক্রিয়ার উপর নির্ভর করে। একটা মানুষ একই আলোক উৎসকে বিভিন্নভাবে দেখতে পারে।

তরঙ্গদৈর্ঘ্য ও বর্ণ শনাক্তকরন
আইজ্যাক নিউটন আবিষ্কাএ করেছিলেন যে সাদা আলোকে যখন কোন প্রিজমের মধ্য দিয়ে পাঠানো হয়, তখন তা বিভিন্ন ভাগে ভাগ হয়ে যায়, যাদের আবার আরেকটি প্রিজমের ভেতর দিয়ে নিয়ে গেলে আবার সাদা আলো পাওয়া যায়।
বর্ণকে উচ্চ থেকে নিম্ন তরঙ্গদৈর্ঘ্যে (বা নিম্ন থেকে উচ্চ কম্পাঙ্কে) অনুযায়ি সাজালে লাল, কমলা, হলুদ, সবুজ, নীল, ও বেগুনী এই ক্রম পাওয়া যায়।তরঙ্গদৈর্ঘ্যে সামান্য পার্থক্যও বর্ণ পাল্টে দিতে পারে; যেমন সবুজাভ নীল ও হলুদের মধ্যে তরঙ্গদৈর্ঘ্যের পার্থক্য হল ১ ন্যানমিটার। যদিও মানুষের চোখ এই পার্থক্য অনেক বড় পার্থক্যেই ধরতে পারে, যখন এই বর্ণালীগত রঙকে একসাথে মেশানো হয়, তখন ক্রোমাটিসিটি অনেক উচ্চ সংখ্যার হতে পারে।
নিম্ন আলোর ক্ষেত্রে দৃষ্টি স্কটপিক হয়। তখন আলো রেটিনার রড কোষ এ ধরা পড়ে।এই কোসগুলো সাধারনত ৫০০ ন্যানমিটার তরঙ্গদৈর্ঘ্যের আলোও ধরতে পারে। উচ্চ আলোর ক্ষেত্রে দৃষ্টি ফোটোপিক হয়। ্তখন আলো রেটিনার কোন কোষ এ ধরা পরে, যা বর্ন দেখার জন্য দায়ী। কোনগুলোও নির্দিষ্ট তরঙ্গদৈর্ঘ্য পর্যন্ত দেখতে পারে, তবে সবচেয়ে বেশী সেন্সেটিভ হয় ৫৫৫ ন্যানোমিটার এর তরঙ্গের দিকে। এই দুটির এলাকার মধ্যে মেসপিক দৃষ্টি কাজ করে এবং সেখানে রদ ও কোন দুটিই সিগন্যাল পাঠায়। এই নিম্ন আলো থেক উচ্চ আলোর প্রত্যক্ষদর্শন একটি পার্থক্যের জন্ম দেয় যাকে পার্কিনজি এফেক্ট বলে।
"সাদা" আলোর উপলব্ধিতা দৃশ্যমান আলোর সন বর্ণের যোগফলে তৈরি হয়। মানুষের ক্ষেত্রে লাল, নীল ও সবুজ ব্যাবহার করে সাদা আলো তৈরী করা যায়, অথবা বিপরীত আলো যেমন নীল ও হলুদ ব্যবহার করেও পাওয়া যায়। [1]
বর্ণ প্রত্যক্ষকরনের দেহতত্ত্ব

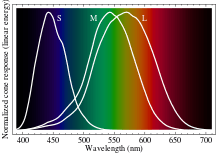
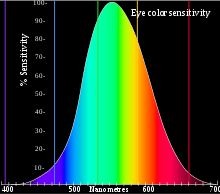
বর্ণ প্রত্যক্ষকরনের ব্যপারটা রেটিনার কিছু কোষ যা বিভিন্ন বর্ণালীগত সংবেদনশীলতার পিগমেন্ট ধারন করে। এদের কোন কোষ বলে। মানুষের ক্ষেত্রে তিন ধরনের কোন কোষ থাকে যা তিনটি আলাদা বর্ণালীর প্রতি সংবেদনশীল, যার ফলে ট্রাইক্রোম্যাটিক বর্ন দৃষ্টি হয়ে থাকে।
প্রত্যেক আলাদা কোন অপসিন এপোপ্রোটিনের তৈরী পিগমেন্ট বহন করে, যা 11-cis-hydroretinal অথবা 11-cis-dehydroretinal দ্বারা যুক্ত থাকে।[2]
কোনগুলো তাদের বর্ণালীগত সংবেদনশীলতার চূড়ার তরঙ্গদৈর্ঘ্য অনুযায়ী বিন্যস্ত থাকেঃ ছোট (S), মাঝারী (M) ও বড় (L) কোন। এই ৩ টি ধরন নির্দিষ্ট বর্ণ অনুযায়ী মিলেনা, যেটা আমরা জেনে এসেছি। এর বদলে, বর্ন প্রত্যক্ষকরন এসব কোষের রেটিনায় পরার ফল হতে শুরু হয় এবং মস্তিষ্কের ভিজুয়াল কর্টেক্স ও এসসোসিয়েটিভ এলাকায় শেষ হয়।
উদাহরনস্বরুপ, যেখানে L কোন লাল রিসেপ্টরের প্রতি সংবেদনশীল, সেখানে মাইক্রোস্পেকট্রোকেমিস্ট্রি দেখায় যে এস্ময় তাদের চুড়ান্ত সংবেদনশীলতা ছিল সবুজাভ হলুদ এলাকায়। S এবং M এর ক্ষেত্রেও এসব দেখা যায় যে এরা সরাসরি নীল ও সবুজ এর প্রতি সংবেদনশীল নয়। তাই বলা যায়, আরজিবি কালার মডেল বর্ণকে বর্নণা করার জন্য ব্যবহৃত হলেও তা মানবচক্ষুর কোনের ধরনের উপর নির্ভর করে বানানো হয়নি।
মানবচক্ষুর কোন কষের পিক রেসপন্স পাল্টায়, সাদাহ্রন বর্ণদৃষ্টিও;[3] আর অন্য কিছু প্রাণীর ক্ষেত্রে এই পলিমরফিক দৃষ্টি অনেক বিশাল হয়।[4]
তত্ত্বসমুহ
বর্ন দর্শনের দুটি পরিপূরক তত্ত্ব হল ট্রাইকোমেট্রিক তত্ত্ব ও অপনেন্ট প্রসেস তত্ত্ব। ১ম তত্ত্বটি দিয়েছিলেন থমাস ইয়াং ও হারম্যান ভন হেমহোল্টয নামক দুই বিজ্ঞানী, ১৯শ শতাব্দীতে, যার ফলে এই তত্ত্বকে ইয়াং-হেমহোল্টয তত্ত্বও বলে। এই তত্ত্ব বলে যে রেটিনার তিন ধরনের কোনগুলো নীল, সবুজ ও লালের প্রতি মাপামাপিভাবেই সংবেদনশীল। আর ২য় তত্ত্বটি দেন ইও্যান হেরিং ১৮৭২ সালে।[5] এতে বলা হয় দৃষ্টিব্যবস্থা বর্ণকে শত্রুর মত দেখেঃ লাল vs. সবুজ, নীল vs. হলুদ, কাল vs. সাদা এমন। দুটো তত্ত্বই সত্য বলে প্রমাণিত, যা দৃষ্টির দেহতত্ত্বের বিভিন্ন ধাপ বর্নণা করে।[5] যেমনভাবে “একটু নেগেটিভ” পজিটিভ নাম্বার বলে কিছু নেই, তেমনি একটি চোখ কোনভাবেই নীলাভ হলুদ বা লালচে সবুজ দেখতে পারবেনা (কিন্তু এমন রং বাইনোকুলার রাইভেলরীর মাধ্যমে দেখা যায়)।
মানুষের চোখের কোন কোষ
| কোন কোষের ধরন | নাম | সীমা | সর্বোচ্চ তরঙ্গদৈর্ঘ্য[6][7] |
|---|---|---|---|
| S | β | ৪০০-৫০০ | ৪২০-৪৪০ |
| M | γ | ৪৫০-৬৩০ | ৫৩৪-৫৫৫ |
| L | ρ | ৫০০-৭০০ | ৫৬৪-৫৮০ |
আলোর তরঙ্গদৈর্ঘ্যের একটি সীমা এসব রিসেপ্টরের ধরনগুলোকে বিভিন্ন কোন অনুযায়ী স্টিমুলেট করে। উদাহরনস্বরুপ হলুদাভ সবুজ L ও M উভয় কোনকেই সমানভাবে স্টিমুলেট করে, কিন্তু S কোনকে হালকাভাবে স্টিমুলেট করে। আবার লাল আলো L কোনকে গাঢ়ভাবে স্টিমুলেট করে, কিন্ত M কে হালকাভাবে করে। S কে বলতে গেলে স্টিমুলেট করেই না। আমাদের মস্তিষ্ক এসব তথ্য রিসেপ্টর থেকে জোগাড় করে যাতে বিভিন্ন তরঙ্গদৈর্ঘ্যের বিভিন্ন আলোকে দেখা যেতে পারে।
অপসিনে যে L ও M থাকে তা আসে X ক্রোমোসোম থেকে্, যাতে খুত থাকলে বর্নান্ধতা হতে পারে। OPN1LW জিন, যা L কোনে অপ্সিন এর জন্যে দায়ী, তা উচ্চ পলিমরফিক হয়।[8] অল্পসংখ্যক মহিলাদের একটি আলাদা বর্ণ রিসেপ্টর থাকতে পারে কারণ তাদের উভয় X ক্রোমসোমে L অপসিনের জিনের জন্য আলাদা অ্যালিল থাকতে পারে। X ক্রোমোসোম অস্বয়ংক্রিয়করন বলতে বুঝায় যে প্রত্যেক কোন সেকে একটিমাত্র অপসিন কাজ করে, এবং কিছু মহিলার এর ফলে টেট্রাক্রোমাটিক বর্ন দৃষ্টি থাকতে পারে।[9] OPN1MW এ শুধু M কোনের অপসিন এর জন্যে দায়ী, তা একদমই বিরল। এবং এতে বর্নালিগত সংবেদনশীলতায় কোন প্রভাব পরেনা।
মানুষের মস্তিষ্কে বর্ণ
বর্ণ প্রক্রিয়াজাতকরন আমাদের ভিজুয়াল সিস্টেমে ইনিশিয়াল কালার অপনেন্ট মেকানিজমের মাধ্যমে খুবই প্রাথমিক লেভেল তৈরী হয়ে যায়। ট্রাইক্রোমেসি এবং অপনেন্ট প্রসেস উভয় তত্ত্বই যদিও সঠিক, কিন্তু ট্রাইক্রোমেসি যেখানে রিসেপ্টরের লেভেলে কাজ করে, অপনেন্ট প্রসেস সেখানে রেটিনাল গ্যাংলিয়ান কোষ এবং তারও পরে কাজ করে। হেরিঙ্গয়ের তত্ত্বে অপনেন্ট মেকানিজম মানে হল বর্ণের বিপ্রতীপ প্রভাব – লাল-সবুজ, নীল-হলুদ, এবং আলো-অন্ধকার। যা-ই হোক, ভিজুয়াল সিস্টেমে, আসলে বিভিন্ন রিসেপ্টরের ধরনটাই বিপরীত হয়। কিছু বামন গ্যাংলিয়ান কোষ L ও M কোন কোষের সক্রিয়তাকে বিপরীত করে, যা লাল-সবুজ বিপ্রতীপ দশার জন্যে দায়ী। কিন্তু আসলে এটি নীল-সবুজ থেকে ম্যাজেন্টার অক্ষ বরাবর কাজ করে। রেটিনার কিছু ছোট বিস্ট্র্যাটিফাইড কোষ S থেকে L ও M এর বিপ্রতীপ দশা তুলে ধরে। যা অনেক সময় নীল-হলুদ বিপ্রতীপ দশার জন্য কাজ করে,কিন্তু আসলে হলুদ-সবুজ থেকে বেগুনী এর অক্ষ বরাবর কাজ করে। আমাদের মস্তিষ্কে এই ভিজুয়াল তথ্য রেটিনার গ্যাংলিয়ান কোষগুলো অপটিক স্নায়ু ও অপটিক শিয়াজমা (একটি পয়েন্ট যেখানে দুটি অপটিক স্নায়ু মিলিত হয় এবং টেমপোরাল ভিজুয়াল ফিল্ডের তথ্য মস্তিষ্কের অপর প্রান্তে পৌছায়) হয়ে আসে। অপটিক শিয়াজিমার পর ভিজুয়াল ট্র্যাক্টগুলো অপটিক ট্র্যাক্টে যায়, যা থ্যালামাস হয়ে ল্যাটারাল জেনিকুলেট নিউক্লিয়াস এ মিলিত হয়।
এই ল্যাটারাল জেনিকুলেট নিউক্লিয়াস লেমিনিতে বিভক্ত হয়। এর আবার ৩ ধরনের হয়ঃ M লেমিনি, যা M কোষ দ্বারা গঠিত। P লেমিনি, যা P কোষ দ্বারা গঠিত। এবং ক্যানিওসেলুলার লেমিনি। M এবং P কোষ রেটিনার L ও M কোন থেকে সমভাবে ইনপুট পায়, যদিও তা ফোভিয়া, কিছু বামন কোষ যা P লেমিনিতে মিলিত হয়। ক্যানিওসেলুলার লেমিনি ছোট বাইস্ট্র্যাটিফাইড গ্যাংলিয়ান কোষ থেকে এক্সন গ্রহণ করে।[10][11]
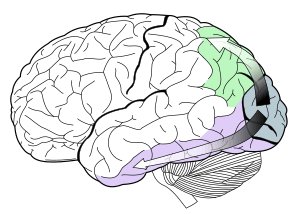
ল্যাটারাল জেনিকুলেট নিউক্লিয়াস এ মিলিত হবার পরে ভিজুয়াল ট্র্যাক্ট আবার প্রাথমিক ভিজুয়াল কর্টেক্স (V1) ( এটি মস্তিষ্কের পেছন দিকে অক্সিপেটাল লোব এ অবস্থিত) এ ফিরে আসে। V1 এর ভেতরে একটি ডিসটিঙ্কট ব্যান্ড (স্ট্রিয়েশন) থাকে। একে আবার “স্ট্রিয়েট কর্টেক্স”ও বলা হয়, অন্যসব কর্তিক্যাল ভিজুয়াল রেজিওনগুলোকে একসাথে “এক্সট্রাস্ট্রিয়েট কর্টেক্স” বলা হয়। এই স্তরে এসে বর্ণ প্রক্রিয়াজাতকরন অনেক জটিল হয়ে পরে।
V1 এ ত্রিবর্ণী পৃথকীকরন ভাঙতে শুরু করে। V1 এর অনেক কোষ বর্ণালির কিছু অংশে অন্যান্য অংশ থেকে ভালভাবে সাড়া দেয়, কিন্তু এই “বর্ণ সুরকরন” অনেক সময় ভিজুয়াল সিস্টেমের অভিযোজন অবস্থার উপর নির্ভর করে আলাদা হতে পারে। একটি কোষ যা উচ্চ তরঙ্গদৈর্ঘ্যের আলোতে ভাল সাড়া দেয়ার কথা যদি আলোটি উজ্জ্বল হয়, আর সব তরঙ্গদৈর্ঘ্যেই সাড়া দিবে যদি আলোটি অনুজ্জ্বল হয়। কারন এসব কোষের বর্ণ সুরকরন স্থায়ী না।। এটা বিশ্বাস করা হয় যে V1 একটি আলাদা ও তুলনামূলক ছোট সংখ্যার নিউরন বর্ন দৃষ্টির জন্য দায়ী। এসব বিশেষজ্ঞ “বর্ণ কোষ”এ অনেক সময় রিসেপ্টিভ ক্ষেত্র থাকে যা স্থানীয় কোন রেশিও ধরতে পারে। এই “ডাবল-অপনেন্ট” কোষ নিগেল ডাউ নামে এক বিজ্ঞানী গল্ডফিশের ক্ষেত্রে প্রমাণ করেন;[12][13] আর তার অস্তিত্ব আছে বলে ব্যাখ্যা দেন ডেভিড হোবেল ও টরস্টেন উইসেল,[14] আর প্রমাণ করেন বেভিল কর্নওয়ে।[15] মার্গারেট লিভিংস্টন ও ডেভিড হোবেল দেখালেন যে ডাবল অপনেন্ট কোষগুলো V1 এ জমা হয়ে থাকে যাদের ব্লবস বলে, এবং এরা জোড়া বর্ণে থাকে, যেমন লাল-সবুজ, ও নীল-হলুদ। লাল-সবুজ কোষগুলো দৃশ্যের একটি পার্টের লাল-সবুজ অংশের সাথে দৃশ্যসংলগ্ন একটি অংশের লাল-সবুজের সাথে তুলনা করে, যা স্থানীয় আলোক বৈসাদৃশ্যতে ভালভাবে সাড়া দেয়। মডেলিং তথ্য জানান দেয় যে ডাবল অপনেন্ট কোষগুলো বর্ণ স্থিতিশীলতার আদর্শ উদাহরন, যার ব্যাখ্যা এডুইন ল্যান্ড তার রেটিন্যাক্স তত্ত্বে দেন।[16]
V1 ব্লবস থেকে বর্ণের তথ্য চলে যায় দ্বিতীয় ভিজুয়াল এরিয়া V2 তে। এর কোষগুলো সবচেয়ে বেশি বর্ন টিউনড অবস্থায় থাকে, আর V1 এর ব্লবসের মত জমা হয়ে পাতলা ডোরাকাটা দাগ তৈরি করে, যে দাগগুলো হয় সাইটোক্রোম অক্সাইডেজ নামক এনজাইম এর জন্য। V2 এর নিউরনগুলো তখন বর্ধিত V4 এর সাথে মিলিত হয়। এই এলাকা শুধু V4 ই নয়, বরং পেছনের দিকের আরো দুটি এলাকা ইনফেরিওর টেমপোরাল কর্টেক্স, V3 এর সামনের দিক, ডর্সালের পেছনে ইনফেরিওর টেমপোরাল কর্টেক্স, এবং TEO এর পেছনের দিকে।[18][19] V4 এর এলাকা বর্ণের প্রতি একচেটিয়াভাবে নিয়োজিত থাকবে এমনটা বলেছলেন সেমির জেকি নামে এক বিজ্ঞানী, কিন্তু তা ভুল প্রমাণিত হয়।[20] বিশেষভাবে V4 এ অরিয়েন্টেশন-সিলেক্টিভ কোষ এর উপস্থিতি বলে যে V4 উভয় বর্ণকেই প্রক্রিয়াজাত করে এবন বর্ণের সাথে সহচর্যে থাকে।[21] V4 এ মিলিমিটার আকারের বর্ণ মডিউল প্রক্রিয়াজাতকরকে গ্লোবস বলে। এটি মস্তিষ্কের সেই প্রথম অংশ যেখানে কালার স্পেস এ পাওয়া বর্ন বর্ণালির সম্পূর্ন সীমায় প্রক্রিয়াজাত করবে।
এনাটমির তথ্যানুযায়ী বর্ধিত V4 এর নিউরনগুলো ইনফেরিওর টেমপোরাল লোব এ প্রবেশ করে। “IT” কর্টেক্স বর্ণের তথ্যগুলোকে আকারে বর্ধিত করে, যদিও এর সঠিক মানদন্ড পাওয়া যায়নি। এমন অস্পষ্টতার পরেও এটি ব্যবহৃত হয় কারণ এটি সঠিক রাস্তা বাৎলে দেয় (V1>V2>V4>IT), যেখাণে ভেন্ট্রাল স্ট্রীম ডর্সাল স্ট্রীম এর চেয়ে আলাদা হয় এবং গতি বুঝতে সাহায্য করা সহ আরো কিছু উপকারী তথ্য দেয়।
বর্ণ প্রত্যক্ষকরনের সাপেক্ষতা
কোনকিছুই নিঃশর্তভাবে বিশাল বর্ণালীর অদৃশ্য অংশ থেকে তড়িৎচৌম্বকীয় বিকিরণের দৃশ্য বর্ণালীকে আলাদা করতে পারেনা। সেদিক থেকে দেখতে বর্ণ পুরোপুরিভাবে তাড়িতচৌম্বকীয় বিকিরণ নয়, বরং একজন দর্শকের দেখার উপলব্ধি মাত্র। অধিকন্তু, আলোর দৃশ্যমান বর্ণালী ও মানব চক্ষুর বর্ণদর্শনের মধ্যে একটা সেচ্ছাচারী ম্যাপিং চলতে থাকে। যদিও প্রায় সবাই-ই এমন ম্যাপিং করতে থাকে, দার্শনিক জন লক দেখলেন যে এর বিকল্পও সম্ভব, এবং একে তিনি “বিপরীত বর্ণালি” নাম দেন, যা একটি চিন্তামূলক পরীক্ষা। উদাহরনস্বরুপ, একজন ব্যক্তি এই বিপরীত বর্ণালির কারণে লালকে সবুজ দেখলেন, এবং সবুজকে লাল দেখলেন। সিনথেশিয়া সাপেক্ষ বর্ন পরীক্ষার এরকমই কিছু কিন্তু উদ্ভাসক উদাহরন যা শুধু আলোই নয়, শব্দ বা আকার দিয়েও ট্রিগার হতে পারে। এভাবে পৃথিবীর ধর্মাবলী থেকে বর্ণ পরীক্ষণের পৃথকীকরনের সম্ভাবনা বলে দেয় যে বর্ণ একটি সাপেক্ষ মনোবিজ্ঞানগত ঘটনা।
হিমবা সম্প্রদায় এর লকের বর্ণকে অন্যান্য ইউরো-আমেরিকানদের চেয়ে আলাদাভাবে বর্ণনা করে এবং সবুজ রঙের ক্লোজ শেডও পৃথক করতে পারে, যা সাধারন মানুষ ধরতে পারেনা।[22] হিমবারা একদমই আলাদা বর্ণসজ্জা তৈরী করেছে যা বর্ণালীকে গাঢ় শেড ( হিমবা ভাষায় জুজু), খুব হালকা ( তাদের ভাষায় ভাপা), উজ্জ্বল নীল এবং সবুজ ( ওদের ভাষায় বুরু) এবং কিছু শুকনো রঙ, যা তাদের জীবনযাপনের ব্যবস্থানুযায়ী তৈরী।
বর্ণ প্রত্যক্ষকরন বস্তুকে কোথায় উপস্থাপন করা হয়েছে সে প্রসঙ্গের অপর অনেকটাই নির্ভর করে। উদাহরনস্বরুপ, নীল, লাল বা বেগুনী আলোর নিচে সাদা কাগজ আমাদের চোখে যথাক্রমে নীল, লাল বা বেগুনী আলোই প্রতিফলন করবে, য়ামাদের মস্তিষ্ক আবার আলোকের প্রভাব পুরন করতে চাইবে এবং উক্ত তিন অবস্থাতেই সাদা কাগজটিকে সাদা ভাবতে বাধ্য করবে। এ ঘটনাকে বর্ণের স্থিতিশীলতা বলে।
অন্যান্য প্রানীতে
অনেক প্রানিই মানুষের “দৃশ্যমান বর্নালি”র বাইরে দেখতে পারে। মৌমাছি এবং আরো অনেক পতঙ্গ অতিবেগুনী রশ্মি দেখতে পারে, যা তাদের ফুলের মধু খুজতে সাহায্য করে। যেসব গাছপালা কীটপতঙ্গের পরাগায়নের উপর নির্ভর করে তা মানুষের দেখা বর্নের চেয়ে অতিবেগুনীতে দেখা বর্ণের উপর নির্ভর করে। পাখিরাও অতিবেগুনী শনাক্ত করতে পারে, এবং কিছু পাখির যৌনমিলনের জন্যেও এই রশ্মি প্রয়োজন।[23][24] যেসব প্রানী অতিবেগুনীর সীমায় দেখতে পারে তারা লাল বা লালের কাছাকাছি তরঙ্গদৈর্ঘ্যের কোন বর্ণ দেখেনা। পাখিরা সামান্য লাল দেখে, কিন্তু তা মানুষের মত না।[25] একটা প্রচলিত ভুল কথা রয়েছে যে গোল্ডফিশ অতিবেগুনী থে অবলোহিত সবই দেখতে পায়,[26] কিন্তু এরা অতিবেগুনী দেখলেও দৃষ্টিসীমা অবলোহিত পর্যন্ত নয়।[27]
এই পার্থক্যের কারন কোন কোষের সংখ্যা ও ধরনে পার্থক্য। স্তন্যপায়ীদের ক্ষেত্রে দুই টাইপ কোন কোষ থাকে, আর এরা লাল-সবুজ বর্নান্ধতাইয় ভোগে। মানুষ অনেক বর সীমায়ই বর্ণ দেখে, কিন্তু তা অন্য স্তন্যপায়ীর সাপেক্ষে মাত্র। বিভিন্ন অস্তন্যপায়ী মেরুদন্ডি প্রানীরা মাউষের মতই দেখতে পারে। আবার কিছু প্রজাতি পাখি, সরীসৃপ, মাছ ও এম্ফিবিয়ানদের ৩ এর চেয়েও বেশী কোন কোষ থাকে, এবং তাদের বর্ণদৃষ্টি মানুষের চেয়েও ভাল।
বেশীরভাগ কাটারহিনির ( বাদর ও শিম্পাঞ্জীদের পূর্বসূরি, মানুষদেরও পূর্বসূরিও ধরা হয় একে) ৩ ধরনের বর্ণ রিসেপ্টর থাকে, যার ফলে ট্রাইক্রোমাটিক বর্ণদৃষ্টি হয়। এসব প্রাইমেটদের ট্রাইক্রোম্যাট বলে। অন্যান্য প্রাইমেট এবং স্তন্যপায়ীরা ডাইক্রোম্যাট হয়, যা দিনের বেলায় স্তন্যপায়ীর ক্ষেত্রে সাধারন দৃষ্টি। নিশাচর স্তন্যপায়ীদের বর্ণান্ধতা অথবা সামান্য বর্ণদৃষ্টি থাকে।ট্রাইক্রোমেট নন-প্রাইমেট স্তন্যপায়ী অনেক বিরল।[28][29]
বিভিন্ন অমেরুদন্ডীর বর্ণদৃষ্টি থাকে। মৌমাছি ও ভ্রমরদের ট্রাইক্রোম্যাটিক বর্ণ দৃষ্টি থাকে যা লালের প্রতি অসংবেদনশীল কিন্তু অতিবেগুনীর দিকে সংবেদনশীল। উদাহরনস্বরুপ “অসমিয়া রুফা”র ট্রাইক্রোম্যাটিক বর্ণ দৃষ্টি থাকে, যা তারা ফুলের পোলেন খুজতে ব্যবহার করে।[30] মাছিদের বর্ণদৃষ্টির প্রয়জনীয়তা নিয়ে কেউ ভাবতে পারে এসব রিসেপ্টর সংবেদনশীলতা তাদের নির্দিষ্ট ভিজুয়াল ইকোলজিকে প্রতিফলিত করবে। যাই হোক, হিমেনোটেরান গ্রুপের কীটপতঙ্গরা ( মাছি, ওয়াস্প ইত্যাদি) ৩ ধরনের ফটোরিসেপ্টর থাকে, যার বর্ণালীগত সংবেদনশীলতা মৌমাছির সমান।[31] পাপিলো প্রজাপতির ৬ ধরনের ফটোরিসেপ্টর থাকে, যাদের সম্ভবত পেন্টাক্রোম্যাটিক দৃষ্টি থাকে।[32] প্রানিজগতের সবচেয়ে জটিল দৃষ্টিব্যবস্থা হল স্টমাটোপডদের, যাদের ১২ টি বর্ণালিগত রিসেপ্টর অনেকগুলো ডাইক্রোম্যাটিক ইউনিট হিসেবে কাজ করে।[33] ট্রপিক্যাল মাছ ও পাখির মত মেরুদন্ডী প্রানীদের অনেকসময় মানুষের চেয়েও জটিল বর্ণ দৃষ্টি ব্যবস্থা থাকতে পারে। এভাবে তারা যে বর্ণগুলো প্রকাশ করে তারা সমপ্রজাতির প্রতি কোন বার্তা পাঠানোর কাজ করে।[34] পাখির দৃষ্টির ক্ষেত্রে চারটি কোন কোষ থেকে টেট্রাক্রোম্যাটিক দৃষ্টি তৈরী হয়। প্রত্যেক কোন প্রধান চারটি ভার্টেব্রাটা কোন ফটোপিগমেন্ট (LWS/ MWS, RH2, SWS2 এবং SWS1) এর একটি ধারন করে এবং এর ভেতরের অংশে বর্ণযুক্ত তৈলবিন্দু আছে। কোনের ভেতরের উজ্জ্বল বর্ণের তৈলবিন্দু কোষের বর্ণালীগত সংবেদনশিলতা কমিয়ে দেয়। ফলে এটা বলা হয় যে কবুতর পেন্টাক্রোম্যাটিক হয়।[35] সরীসৃপ ও এম্ফিবিয়ানদেরও চার ধরনের কোন থাকে ( কখনও ৫ টি) এবং এরা প্রায় মানুষের মতই দেখে।, অথবা তার চেয়েও বেশী। আরও বলা যায়, নিশাচর গেকো অনুজ্জ্বল আলোতেও দেখতে পারে।[36]
স্তন্যপায়ীদের বিবর্তনের সাথে সাথে বর্ণদৃষ্টির অংশ হারিয়ে যায়, আবার কিছুর ক্ষেত্রে সেটা ফিরে আসে জিন ডুপ্লিকেশন এর মাধ্যমে। ইউথারিয়ান স্তন্যপায়ী অন্যান্য প্রাইমেটের চেয়ে কম প্রভাবশালী দুটি রিসেপ্টরের বর্ণ প্রত্যক্ষদর্শিতা নিয়ে চলছে, যা কেবল হলুদ, সবুজ ও নীলের পার্থক্য জানে কিন্তু লাল ও কমলার পার্থক্য জানেনা। এমন কিছু প্রমাণও আছে যে কিছু স্তন্যপায়ি, যেমন বিড়ালের উচ্চ তরঙ্গদৈর্ঘ্যের পার্থক্য ধরার ক্ষমতা আছে, তাদের অপসিন জিনে এমিনো এসিডের পরিব্যক্তির মাধ্যমে।[37] লাল দেখার ক্ষমতা প্রাইমেট স্তন্যপায়ীদের জন্য দরকারী ছিল, যা অন্তত ফলের পার্থক্য শেখায়। যাই হোক, প্রাইমেটদের মধ্যেও বর্তমান ও পুরাতন বাদরদের মধ্যে বর্ণদৃষ্টির পার্থক্য বিদ্যমান। বর্তমান বাদরদের এই লেভেলের বর্ণ সংবেদনশিলতা থাকতেও পারে নাও থাকতে পারেঃ বেশীরভাগ প্রজাতীর ক্ষেত্রে পুরুষ ডাইক্রোম্যাট হয়, আর ৬০% নারী ট্রাইক্রোম্যাট হয়। কিন্তু কিছু বাদর মনোক্রোম্যাটা হয়। আর হাওলার বাদর ট্রাইক্রোম্যাট হয়।[38][39][40][41] হলুদ-সবুজ সংবেদনশীল অপসিন প্রোটিনের ( যার ফলে লাল থেকে সবুজ আলাদা করা যায়) জিনের জন্যে পুরুষ ও নারী প্রজাতিতে ভিজুয়াল সংবেদনশীলতার পার্থক্য থাকে X ক্রোমোসমের ভিত্তিতে।
কিছু মারসুপিয়াল যেমন মোটা লেজের ডানার্ট (Sminthopsis Crassicaudata) ট্রাইক্রোম্যাটিক বর্নদৃষ্টি দেখায়।[42]
সামুদ্রিক স্তন্যপায়ী যাদের নিম্ন আলোতে দেখার ভাল ক্ষমতা আছে, তারা মনোক্রোম্যাটিক হয়।
| অবস্থা | কোন কোষের ধরন | বর্ণ পারসিভ করার সংখ্যা | বাহক |
|---|---|---|---|
| মনোক্রোমেসি | ১ | ১০০ | সামুদ্রিক স্তন্যপায়ী,বাদর , অসট্রেলিয়ান সি লায়ন, এক্রোমেট প্রাইমেট |
| ডাইক্রোমেসি | ২ | ১০০০০ | নন-প্রাইমেট স্তন্যপায়ী, বর্ণান্ধ প্রাইমেট |
| ট্রাইক্রোমেসি | ৩ | ১০ মিলিয়ন[43] | বেশীরভাগ প্রাইমেট, উচ্চতর শিম্পাঞ্জী গোত্র, মাসুপিয়াল, কিছু পোকা (মৌমাছি) |
| টেট্রাক্রোমেসি | ৪ | ১০০ মিলিয়ন | বেশীরভাগ সরীসৃপ, এম্ফিবিয়া গোত্রের প্রানী, পাখি এবং কীট |
| পেন্টাক্রোমেসি | ৫ | ১০ বিলিয়ন | কিছু পোকা, কিছু পাখি (কবুতর) |
বিবর্তন
বর্ণ প্রত্যক্ষকরন ব্যবস্থা বিবর্তনের উপর অনেকটাই নির্ভর করে, যার মধ্যে সবচেয়ে গুরুত্বপূর্ন ধরা যায় সন্তোষজনকভাবে খাদ্যের উৎস চেনা। তৃনভোজী প্রাইমেটদের ক্ষেত্রে বর্ণ প্রত্যক্ষকরন হল ঠিকভাবে পাতা চেনা। হামিংবার্ডএর ক্ষেত্রে সেটা আবার নির্দিষ্ট ফুল চেনা। অপরপক্ষে, নিশাচর প্রানীর ক্ষেত্রে বর্ণদৃষ্টি অনেকটা অনুন্নত, কারণ কোনগুলো কাজ করতে পর্যাপ্ত আলো প্রয়োজন। এখন প্রমাণ পাওয়া গেছে যে অতিবেগুনী রশ্মি প্রাণীজগৎএর বিভিন্ন শাখায় বর্ণ প্রত্যক্ষকরনে অংশ নেয়, বিশেষত কীটপতঙ্গের ক্ষেত্রে। সাধারণভাবে, অপ্টিকাল বর্ণালী বস্তুর ইলেকট্রনিক ট্রান্সিশানকে বেষ্টন করে রাখে এবং পরিবেশের ব্যাপারে তথ্য জোগাড়ে সাহায্য করে।
প্রাইমেটদের ট্রাইক্রোম্যাটিক বর্ণদৃষ্টি বিবর্তিত হয়ে উত্তরসূরি বর্তমান বাদর, শিম্পাঞ্জী, এবং মানুষে ডায়ার্নাল সক্রিয়তায় পরিণত হয়েছে এবং গাছের ফল ও ফুল খেতে শুরু করে।[44] অতিবেগুনীর পার্থক্যসহ বর্ণদৃষ্টি কিছু সংখ্যক এন্থ্রোপড এ উপস্থিত আছে, যা মেরুদন্ডীডের বাইরে একমাত্র উদাহরন।[45]
কিছু প্রানী অতিবেগুনী বর্ণালীতে বর্ণের পার্থক্য করতে জানে। এই অতিবেগুনী বর্ণালী মানুষের দৃষ্টিসীমার বাইরে কিছু কাটারেক্ট সার্জারীর রোগী বাদে।[46] পাখি, কচ্ছপ, গিরগিটি, কিছু মাছ, এবং কিছু রডেন্টের অতিবেগুনী রশ্মি দেখার ক্ষমতা আছে।[47] তারা তাদের খাদ্য ও স্বভাবিক জীবনে অতিবেগুনী রশ্মি দেখতে পারে, আ মানুষের কাছে অদৃশ্য।
পাখিদের কাছে এই দৃষ্ট অত্যন্ত গুরুত্বপূর্ণ। এর ফলে পাখি খুব স্বল্প দূরত্বের শিকার দেখতে পারে এবং শিকারীর কাছ থেকে পালাতেও পারে, এমনকি উচ্চগতিতে থাকা সত্ত্বেও। তারা এই অতিবেগুনী রশ্মি ব্যবহার করে তাদের সঙ্গীনীকেউ চিনতে পারে।[48][49]
বর্ণ প্রত্যক্ষকরনের গণিত
একটি বাস্তব বর্ণ কিছু নিখাদ বর্ণালীগত বর্ণ এর সমষ্টিমাত্র। যেহেতু তত্ত্বানুযায়ি অসংখ্য বর্ণালিগত বর্ণ রয়েছে, তাই সকল বাস্তব বর্ণের সেটকে অসীম মাত্রার ভেক্টর স্পেস, আরও ভালভাবে বললে হিলবার্ট স্পেস ভাবা যায়।আমরা এই স্পেসকে Hcolor বলি। প্রায়োগিকভাবে, বাস্তব বর্ণগুলোকে সিমপ্লেক্স এর কোন ভাবা যায় (গাণিতিকভাবে), যাদের ছেদবিন্দুতে বর্ণালীগত বর্ণ, ভরকেন্দ্রে সাদা থাকে এবং চূড়ায় কাল থাকে, এবং মনোক্রোম্যাটিক বর্ণগুলো ছেদবিন্দু ও চূড়ার মধ্যবর্তী কোন স্থানে একই লাইন বরাবর বিরাজমান থাকে।
Hcolor এর একটি এলিমেন্ট C যা দৃশ্যমান তরঙ্গদৈর্ঘ্যের সীমা (যা বাস্তব সংখ্যার মধ্যবর্তী পার্থক্য হয় [Wmin, Wmax] থেকে বাস্তব সংখ্যার ফাংশান, যা [Wmin, Wmax] প্রত্যেক তরঙ্গদৈর্ঘ্য W কে এর তীব্রতা C(w) তে আরোপন করে।
একটি মানবদৃষ্ট বর্ণকে ৩টি সংখ্যর মডেল হিসেবে ভাবা যায় যে সীমায় ৩টি কোনই স্টিমুলেট করা হবে। এভাবে একটি মানবদৃষ্ট বর্ণকে তৃতীয় মাত্রার ইউক্লীডীয় স্পেস এর একটি বিন্দু ভাবা যায়। আমরা এই স্পেসকে R3color বলি।
যেহেতু প্রত্যেক তরঙ্গদৈর্ঘ্য w তিনটি কোন কোষকে একটি জানা সীমায় স্টিমুলেট করে, সেহেতু এই তিনটি সীমাকে তাদের কোন কোশের নাম S, M, L অনুযায়ী ৩টি ফাংশান যথাঃ s(w), m(w), l(w) নাম দেয়া যায়।
একটি আলোক বীম যেহেতু অনেক তরঙ্গদৈর্ঘ্যে দ্বারা গঠিত হতে পারে, সেহেতু যে সীমায় Hcolor এ বাস্তব বর্ণ C প্রত্যেক কোন কোষকে স্টিমুলেট করবে সেটা নির্নয় করতে গেলে আমাদের C(w)*s(w), C(w)*m(w), এবং C(w)*l(w) এর যোগজীকরন করতে হবে, যার ব্যপ্তি হবে [Wmin, Wmax]। ফলাফলকে ৩ দিয়ে গুন করলে আমরা যা পাই তা প্রত্যেক বাস্তব বর্ণ C এর নির্দিষ্ট প্রত্যক্ষদর্শিত বর্ণ এর সাথে মিলে যায়। এই মিলিত হওয়াটাকে সহজভাবে রৈখিক ভাবেই পাওয়া গেছে। এটাও সহজভাবে দেখা যায় যে বাস্তব স্পেস Hcolor এর বিভিন্ন এলিমেন্ট R3color এর একই প্রত্যক্ষদর্শিত বর্ণেই পাওয়া যায়, তাই একটি প্রত্যক্ষদর্শিত বর্ণ একতি বাস্তব বর্ণের কাছে অনন্য নাও হতে পারে।
এভাবে মানবদৃষ্ট বর্ণ প্রত্যক্ষকরণকে অসীম মাত্রার হিলবার্ট স্পেস Hcolor থেকে তিন মাত্রার ইউক্লীডীয় স্পেস R3color পর্যন্ত একটি নির্দিষ্ট, অস্বতন্ত্র, রৈখিক ম্যাপিং দ্বারা পরিমাপ করা হয়।
প্রায়োগিকভাবে, রৈখিক ম্যাপিংয়ের মাধ্যমে সিমপ্লেক্সের উপরে কোন যার ছেদবিন্দু বর্ণালীগত বর্ণ, তার ছবি R3color এরও একটি কোন হবে। ছেদবিন্দু থেকে কোনের বাইরে যেতে থাকলে তা একই ক্রোমাটিসিটি তে বাড়ন্ত তিব্রতা নির্দেশ করে। এই কোনের শুধু প্রস্থ নিলে তা একটি দুই মাত্রার ক্রোমাটিসিটি স্পেস তৈরী করে। এই তিন মাত্রার কোন আর এই দুই মাত্রার কোন উভয়েই উত্তল অংশের সেট, যার মানে হল বর্ণালীর মিশ্রনও একটি বর্ণ।

বাস্তবে, কোন একটি ব্যক্তি বা বস্তুর বিভিন্ন বাস্তব বর্ণ স্টিমুলাই এর প্রতি তিনটি কোনের সাড়া দেয়ার মাত্রা পরিমাপ করা অনেক কঠিন। এর পরিবর্তে মনোবিজ্ঞানগতভাবে এ মাপ নেয়া হয়। সাধারণত তিনটি টেস্ট লাইট ব্যবহৃত হয়, যাদের আমরা S, M, L বলতে পারি। মানুষের প্রত্যক্ষদর্শনের স্পেসের ক্রমাঙ্ক নির্ণয়ের জন্যে বিজ্ঞানীরা কিছু লোককে S, M, L এর জন্যে কিছু নির্দিষ্ট কম্বিনেশন ঘুরিয়ে তীব্রতা(IS, IM, IL) পাল্টিয়ে যেকোন বর্ন তৈরী করতে বলেন, যতক্ষন কোন মিল পাওয়া না যায়। জেনে রাখা ভাল বাস্তবে S,M,L এর যেকোন একটিকে বাস্তব বর্ণ পরীক্ষনে কিছু তীব্রতায় যোগ করা হয়, এবং সেই কম্বিনেশন তৈরি হয় বাকী দুটি লাইটের রৈখিক কম্বিনেশন দ্বারা, এবং প্রায় যথাযথ ফলাফল পাওয়া যায়।
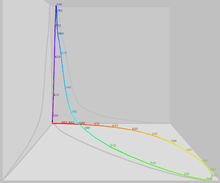
ফলাফলে প্রাপ্ত তীব্রতার কম্বিনেশনগুলোকে ৩ স্পেসের একটি সাবসেট ভাবলে মানুষের প্রত্যক্ষদর্শিত বর্ণ স্পেসের একটি মডেল তৈরী হয় (জানা ভাল যে যখন S,M,L এর একটি টেস্ট কালারে যোগ করা হয়, তখন এর তীব্রতাকে ঋনাত্মক ধরা হয়।)। আবার এটি গানিতীকভাবে একটি কোন হয়, চতুর্ভূজ না, কিন্তু তবুও উৎস যে ৩ মাত্রার স্পেস থেকে উৎপন্ন হয় তার সব রশ্মি একটি উত্তল সেট তৈরী করে। আবার, এই কোনের একটি ধর্ম আছে যার ফলে এটি উৎস থেকে তিব্রতা অনুযায়ি আনুপাতিকভাবে দূরে সরতে থাকে। আবার কোনটির প্রস্থ একটি সমান আকারে থাকে, যার স্পেসের “ক্রোমাটিসিটি”; যা আবার CIE 1931 Color Space এর X+Y+Z এর ধ্রুবতার জন্য দায়ি, এবং এটি থেকে CIE ক্রোমাটিসিটি ডায়াগ্রাম আসে।
এই ব্যবস্থা ধারণা করে যে যেকোন বর্ণ অথবা বর্ণালীর বাইরের বর্ণ যা ক্রোমাটিসিটি ডায়াগ্রামের বাইরে তাদের জন্য অসীম সংখ্যক বিচ্ছিন্ন বাস্তব বর্ণালী রয়েছে যা অন্যান্য বর্ণের মতোই প্রত্যক্ষদর্শী। তাই সাধারণভাবে আমরা যে বর্ণালীগত বর্ণের কথা উপলব্ধি করি তাদের নির্দিষ্ট কম্বিনেশন বলে কিছুই নেই, বরং সেখানে বর্ণ্টি তৈরী হবার এমন অসংখ্য সম্ভাবনা আছে। সীমার মধ্যে যেসব নিখাদ বর্ণালীগত বর্ণ আছে তাদের কেবল মিলিত তরঙ্গদৈর্ঘ্যের আলোতেই প্রত্যক্ষ করা সম্ভব, যখন বেগুনী বর্ণের লাইনে থাকা বর্ণের সীমা দৃশ্যমান বর্ণালীতে নিখাদ বেগুনী ও নিখাদ লালের নির্দিষ্ট অনুপাত থেকেই বের করা যায়।
CIE ক্রোমাটিসিটি ডায়াগ্রাম অশ্বক্ষুড়াকৃতির হয়, যার বাকা প্রান্ত সকল বর্ণালীগত বর্নের জন্য দায়ী। এবং বাদবাকী সোজা প্রান্ত পরিপৃক্ত রক্তবর্ণ এর জন্য দায়ী, যা লাল ও বেগুনীর মিশ্রনে তৈরী।
ক্রোমাটিক অভিযোজন
বর্ণবিজ্ঞানে ক্রোমাটিক অভিযোজন হল একই বস্তুকে দুটি আলাদা উৎসের কাছে রেখে তাকে বর্ণনা করা। এর ব্যবহার হচ্ছে ক্রোমাটিক অভিযোজন ট্রান্সফর্ম যা নিউট্রাল বস্তুর নিউট্রাল থাকা রেকর্ড করবে, অন্য বর্ণগুলো বাস্তবধর্মী রেখে।[50] উদাহরণস্বরুপ, ক্রোমাটিক অভিযোজন ট্রান্সফর্ম ব্যবহৃত হয় যখন ICC প্রোফাইল এর সাথে বিভিন্ন শ্বেত পয়েন্ট এর ছবি কনভার্ট করা হয়।[51] উদাহরনস্বরুপ, এডবি ফটোশপ ব্রাডফোর্ড CAT ব্যবহার করে। বর্ণদৃষ্টিতে ক্রোমাটিক অভিযোজন ট্রান্সফর্ম বর্ণের স্থিতিশীলতাকে নির্দেশ করে; যা ভিজুয়াল ব্যবস্থার এমন এক ক্ষমতা যার ফলে একটি বস্তুর বাহ্যিক রূপকে একটি বিশাল আলোক উৎসের সীমায় সংরক্ষন করে রাখে।[52]
আরো দেখুন
| উইকিমিডিয়া কমন্সে বর্ণের দৃশ্যমানতা সংক্রান্ত মিডিয়া রয়েছে। |
- বর্ণান্ধতা
- বর্ণতত্ত্ব
- বিপরীত বর্ণালী
- মৌলিক বর্ণ
- প্রত্যক্ষদর্শন
Hcolor এর একটি এলিমেন্ট C যা দৃশ্যমান তরঙ্গদৈর্ঘ্যের সীমা (যা বাস্তব সংখ্যার মধ্যবর্তী পার্থক্য হয় [Wmin, Wmax] থেকে বাস্তব সংখ্যার ফাংশান, যা [Wmin, Wmax] প্রত্যেক তরঙ্গদৈর্ঘ্য W কে এর তীব্রতা C(w) তে আরোপন করে।
References
- "Eye, human." Encyclopædia Britannica 2006 Ultimate Reference Suite DVD, 2009.
- Nathans, Jeremy; Thomas, Darcy; Hogness, David S. (এপ্রিল ১১, ১৯৮৬)। "Molecular Genetics of Human Color Vision: The Genes Encoding Blue, Green, and Red Pigments"। Science। 232 (4747): 193–202। doi:10.1126/science.2937147। PMID 2937147। জেস্টোর 169687। বিবকোড:1986Sci...232..193N।
- Neitz J, Jacobs GH (১৯৮৬)। "Polymorphism of the long-wavelength cone in normal human color vision"। Nature। 323 (6089): 623–5। doi:10.1038/323623a0। PMID 3773989। বিবকোড:1986Natur.323..623N।
- Jacobs GH (জানুয়ারি ১৯৯৬)। "Primate photopigments and primate color vision"। Proc. Natl. Acad. Sci. U.S.A.। 93 (2): 577–81। doi:10.1073/pnas.93.2.577। PMID 8570598। পিএমসি 40094

- Hering, Ewald (১৮৭২)। "Zur Lehre vom Lichtsinne"। Sitzungsberichte der Mathematisch–Naturwissenschaftliche Classe der Kaiserlichen Akademie der Wissenschaften। K.-K. Hof- und Staatsdruckerei in Commission bei C. Gerold's Sohn। LXVI. Band (III Abtheilung)।
- Wyszecki, Günther; Stiles, W.S. (১৯৮২)। Color Science: Concepts and Methods, Quantitative Data and Formulae (2nd সংস্করণ)। New York: Wiley Series in Pure and Applied Optics। আইএসবিএন 0-471-02106-7।
- R. W. G. Hunt (২০০৪)। The Reproduction of Colour (6th সংস্করণ)। Chichester UK: Wiley–IS&T Series in Imaging Science and Technology। পৃষ্ঠা 11–2। আইএসবিএন 0-470-02425-9।
- Verrelli BC, Tishkoff SA (সেপ্টেম্বর ২০০৪)। "Signatures of Selection and Gene Conversion Associated with Human Color Vision Variation"। Am. J. Hum. Genet.। 75 (3): 363–75। doi:10.1086/423287। PMID 15252758। পিএমসি 1182016
- Roth, Mark (2006). "Some women may see 100 million colors, thanks to their genes" Post-Gazette.com
- R.W. Rodieck, "The First Steps in Seeing". Sinauer Associates, Inc., Sunderland, Massachusetts, USA, 1998.
- Hendry, Stewart H. C.; Reid, R. Clay (১৯৭০-০১-০১)। "SH Hendry, RC Reid, "The Koniocellular Pathway in Primate Vision". Annual Reviews Neuroscience, 2000, vol. 23, pp. 127-53"। Annual Review of Neuroscience। Annualreviews.org। 23: 127–53। doi:10.1146/annurev.neuro.23.1.127। PMID 10845061। সংগ্রহের তারিখ ২০১২-০৯-০৯।
- Nigel W. Daw (১৭ নভেম্বর ১৯৬৭)। "Goldfish Retina: Organization for Simultaneous Color Contrast"। Science। 158 (3803): 942–4। doi:10.1126/science.158.3803.942। PMID 6054169। বিবকোড:1967Sci...158..942D।
- Bevil R. Conway (২০০২)। their Neural Mechanisms of Color Vision: Double-Opponent Cells in the Visual Cortex। Springer। আইএসবিএন 1-4020-7092-6।
- Conway BR (১৫ এপ্রিল ২০০১)। "Spatial structure of cone inputs to color cells in alert macaque primary visual cortex (V-1)"। J. Neurosci.। 21 (8): 2768–83। PMID 11306629।
- John E. Dowling (২০০১)। Neurons and Networks: An Introduction to Behavioral Neuroscience। Harvard University Press। আইএসবিএন 0-674-00462-0।
- McCann, M., ed. 1993. Edwin H. Land's Essays. Springfield, Va.: Society for Imaging Science and Technology.
- Judd, Deane B.; Wyszecki, Günter (১৯৭৫)। Color in Business, Science and Industry। Wiley Series in Pure and Applied Optics (third সংস্করণ)। New York: Wiley-Interscience। পৃষ্ঠা 388। আইএসবিএন 0-471-45212-2।
- Conway BR, Moeller S, Tsao DY (২০০৭)। "Specialized color modules in macaque extrastriate cortex"। Neuron। 56 (3): 560–73। doi:10.1016/j.neuron.2007.10.008। PMID 17988638।
- Conway BR, Tsao DY (২০০৯)। "Color-tuned neurons are spatially clustered according to color preference within alert macaque posterior inferior temporal cortex"। Proc Natl Acad Sci U S A। 106 (42): 18035–18039। doi:10.1073/pnas.0810943106। PMID 19805195। পিএমসি 2764907
- John Allman; Steven W. Zucker (১৯৯৩)। "On cytochrome oxidase blobs in visual cortex"। Laurence Harris; Michael Jenkin। Spatial Vision in Humans and Robots: The Proceedings of the 1991 York Conference। Cambridge University Press। আইএসবিএন 0-521-43071-2।
- Zeki S (২০০৫)। "The Ferrier Lecture 1995 Behind the Seen: The functional specialization of the brain in space and time"। Philosophical Transactions of the Royal Society B। 360 (1458): 1145–1183। doi:10.1098/rstb.2005.1666। PMID 16147515। পিএমসি 1609195
- Roberson, Davidoff, Davies & Shapiro. referred by Debi Roberson, University of Essex 2011
- Cuthill, Innes C (১৯৯৭)। "Ultraviolet vision in birds"। Peter J.B. Slater। Advances in the Study of Behavior। 29। Oxford, England: Academic Press। পৃষ্ঠা 161। আইএসবিএন 978-0-12-004529-7।
- Jamieson, Barrie G. M. (২০০৭)। Reproductive Biology and Phylogeny of Birds। Charlottesville VA: University of Virginia। পৃষ্ঠা 128। আইএসবিএন 1-57808-386-9।
- Varela, F. J.; Palacios, A. G.; Goldsmith T. M. "Color vision of birds" in Ziegler & Bischof (1993) 77–94
- "True or False? "The common goldfish is the only animal that can see both infra-red and ultra-violet light." - Skeptive"। ডিসেম্বর ২৪, ২০১৩ তারিখে মূল থেকে আর্কাইভ করা। সংগ্রহের তারিখ সেপ্টেম্বর ২৮, ২০১৩।
- Neumeyer, Christa (২০১২)। "Chapter 2: Color Vision in Goldfish and Other Vertebrates"। Lazareva, Olga; Shimizu, Toru; Wasserman, Edward। How Animals See the World: Comparative Behavior, Biology, and Evolution of Vision। Oxford Scholarship Online। আইএসবিএন 978-0-195-33465-4।
- Ali, Mohamed Ather; Klyne, M.A. (১৯৮৫)। Vision in Vertebrates। New York: Plenum Press। পৃষ্ঠা 174–175। আইএসবিএন 0-306-42065-1।
- Jacobs, G. H. (১৯৯৩)। "The Distribution and Nature of Colour Vision Among the Mammals"। Biological Reviews। 68 (3): 413–471। doi:10.1111/j.1469-185X.1993.tb00738.x। PMID 8347768।
- Menzel, R.; Steinmann, E.; Souza, J. De; Backhaus, W. (১৯৮৮-০৫-০১)। "Spectral Sensitivity of Photoreceptors and Colour Vision in the Solitary Bee, Osmia Rufa"। Journal of Experimental Biology। 136 (1): 35–52। আইএসএসএন 0022-0949।
- Osorio D, Vorobyev M (জুন ২০০৮)। "A review of the evolution of animal colour vision and visual communication signals"। Vision Research। 48 (20): 2042–2051। doi:10.1016/j.visres.2008.06.018। PMID 18627773।
- Arikawa K (নভেম্বর ২০০৩)। "Spectral organization of the eye of a butterfly, Papilio"। J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol.। 189 (11): 791–800। doi:10.1007/s00359-003-0454-7। PMID 14520495।
- Cronin TW, Marshall NJ (১৯৮৯)। "A retina with at least ten spectral types of photoreceptors in a mantis shrimp"। Nature। 339 (6220): 137–40। doi:10.1038/339137a0। বিবকোড:1989Natur.339..137C।
- Kelber A, Vorobyev M, Osorio D (ফেব্রুয়ারি ২০০৩)। "Animal color vision—behavioural tests and physiological concepts"। Biol Rev Camb Philos Soc। 78 (1): 81–118। doi:10.1017/S1464793102005985। PMID 12620062।
- Introducing Comparative Colour Vision Colour Vision: A Study in Cognitive Science and the Philosophy of Perception, By Evan Thompson
- Roth, Lina S. V.; Lundström, Linda; Kelber, Almut; Kröger, Ronald H. H.; Unsbo, Peter (মার্চ ৩০, ২০০৯)। "The pupils and optical systems of gecko eyes"। Journal of Vision। 9 (3:27): 1–11। doi:10.1167/9.3.27। PMID 19757966।
- Shozo Yokoyamaa and F. Bernhard Radlwimmera, "The Molecular Genetics of Red and Green Color Vision in Mammals", Genetics, Vol. 153, 919–932, October 1999.
- Jacobs G. H.; Deegan J. F. (২০০১)। "Photopigments and color vision in New World monkeys from the family Atelidae"। Proceedings of the Royal Society B: Biological Sciences। 268 (1468): 695–702। doi:10.1098/rspb.2000.1421।
- Jacobs G. H., Deegan J. F., Neitz, Neitz J., Crognale M. A. (১৯৯৩)। "Photopigments and color vision in the nocturnal monkey, Aotus"। Vision Research। 33 (13): 1773–1783। doi:10.1016/0042-6989(93)90168-V। PMID 8266633।
- Mollon J. D.; Bowmaker J. K.; Jacobs G. H. (১৯৮৪)। "Variations of color vision in a New World primate can be explained by polymorphism of retinal photopigments"। Proceedings of the Royal Society B: Biological Sciences। 222 (1228): 373–399। doi:10.1098/rspb.1984.0071। বিবকোড:1984RSPSB.222..373M।
- Sternberg, Robert J. (2006): Cognitive Psychology. 4th Ed. Thomson Wadsworth.
- Arrese CA, Beazley LD, Neumeyer C (মার্চ ২০০৬)। "Behavioural evidence for marsupial trichromacy"। Curr. Biol.। 16 (6): R193–4। doi:10.1016/j.cub.2006.02.036। PMID 16546067।
- Judd, Deane B.; Wyszecki, Günter (১৯৭৫)। Color in Business, Science and Industry। Wiley Series in Pure and Applied Optics (3rd সংস্করণ)। New York: Wiley-Interscience। পৃষ্ঠা 388। আইএসবিএন 0-471-45212-2।
- Pinker, Steven (১৯৯৭)। How the Mind Works। New York: Norton। পৃষ্ঠা 191। আইএসবিএন 0-393-04535-8।
- Koyanagi, M.; Nagata, T.; Katoh, K.; Yamashita, S.; Tokunaga, F. (২০০৮)। "Molecular Evolution of Arthropod Color Vision Deduced from Multiple Opsin Genes of Jumping Spiders"। Journal of Molecular Evolution। 66 (2): 130–137। doi:10.1007/s00239-008-9065-9। PMID 18217181।
- David Hambling (মে ৩০, ২০০২)। "Let the light shine in: You don't have to come from another planet to see ultraviolet light"। EducationGuardian.co.uk।
- Jacobs GH, Neitz J, Deegan JF (১৯৯১)। "Retinal receptors in rodents maximally sensitive to ultraviolet light"। Nature। 353 (6345): 655–6। doi:10.1038/353655a0। PMID 1922382। বিবকোড:1991Natur.353..655J।
- FJ Varela; AG Palacios; TM Goldsmith (১৯৯৩)। Bischof, Hans-Joachim; Zeigler, H. Philip, সম্পাদকগণ। Vision, brain, and behavior in birds। Cambridge, Mass: MIT Press। পৃষ্ঠা 77–94। আইএসবিএন 0-262-24036-X।
- IC Cuthill; JC Partridge; ATD Bennett; SC Church; NS Hart; S Hunt (২০০০)। "Ultraviolet Vision in Birds"। Advances in the Study of Behavior। 29। পৃষ্ঠা 159–214।
- Süsstrunk, Sabine. Chromatic Adaptation
- Lindbloom, Bruce. Chromatic Adaptation
- Fairchild, Mark D. (২০০৫)। "8. Chromatic Adaptation"। Color Appearance Models। Wiley। পৃষ্ঠা 146। আইএসবিএন 0-470-01216-1।
External links
- Feynman's lecture on color vision
- Peter Gouras, "Color Vision", Webvision, University of Utah School of Medicine, May 2009.
- James T. Fulton, "The Human is a Blocked Tetrachromat", Neural Concepts, July 2009.
- Vurdlak, "Mega Color Blindness Test", Mighty Optical Illusions, March 2009.
- Clive (Max) Maxfield, "Color Vision: One of Nature's Wonders", CliveMaxfield.com, 2006.
- Egopont, "Color Vision Test".
- Lintonapps, "Color Vision Test for Iphone"
- Bruce McEvoy (২০০৮)। "Color vision"। সংগ্রহের তারিখ ২০১২-০৩-৩০।
- What colors do animals see? Web Exhibits
- The Science of Why No One Agrees on the Color of This Dress