RIG-I-like receptor
RIG-I-like receptors (retinoic acid-inducible gene-I-like receptors, RLRs) are a type of intracellular pattern recognition receptor involved in the recognition of viruses by the innate immune system.[1][2] RIG-I (retinoic-acid inducible gene I, also known as DDX58) is the best characterized receptor within the RIG-I like receptor (RLR) family. Together with MDA5 (melanoma differentiation-associated 5) and LGP2 (laboratory of genetics and physiology 2), this family of cytoplasmic pattern recognition receptors (PRRs) are sentinels for intracellular viral RNA that is a product of viral infection. The RLR receptors provide frontline defence against viral infections in most tissues.
RLR ligands
The RIG-I receptor prefers to bind short (<2000 bp) single- or double-stranded RNA carrying an uncapped 5’ triphosphate and additional motifs such as poly-uridine rich RNA motifs.[3][4] RIG-I triggers an immune response to RNA viruses from various families including the paramyxoviruses (e.g. measles), rhabdoviruses (e.g. vesicular stomatitis virus) and orthomyxoviruses (e.g. influenza A).[5][6][7][8][9] MDA5 ligands are poorly characterized, but the preference is for long double-stranded RNA (>2000 bp), such as the replicative form of picornavirus RNA that is found in picornavirus-infected cells.[10][11] LGP2 binds to blunt-ended double-stranded RNA of variable length,[12][13] and also to RNA-bound MDA5 to regulate filament formation.[14] The latter is linked to LGP2’s recognition of picornaviruses (e.g. encephalomyocarditis virus), as per MDA5.[15]
Structural features
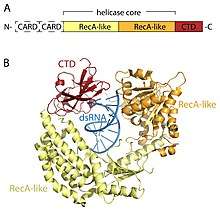
The RLR receptors are members of the DEAD-box (SF2) helicase family (despite containing a DExD/H motif, rather than the DEAD motif characteristic of the family) and share a common domain architecture. All contain a catalytic helicase core made up of two RecA-like domains. The catalytic helicase core contains at least 9 highly conserved sequence motifs that coordinate ATP and RNA binding and the hydrolysis of ATP to unwind RNA. A C-terminal domain (CTD; InterPro: IPR021673) follows the helicase core and this domain also binds viral RNA. Distinct RNA-binding loops within the CTD of the three RLRs dictate the type of RNA that they can bind.[16] In addition to the helicase core and CTD, RIG-I and MDA5 have two N-terminal CARD (caspase active recruitment domains) that are essential to the initiation of downstream signaling. LGP2 is dissimilar to both RIG-I and MDA5 as it lacks the CARD signaling domains and instead is implicated as a positive and negative regulator of RIG-I and MDA5.[17][18][19][20][21][22][23]
Activation of signaling
In uninfected cells that are absent of viral RNA RIG-I exists in an inactive conformation in which the CARD domains are masked due to their interaction with the CTD.[24] Upon binding RNA, RIG-I changes into a conformation in which the CARD domains are exposed and ‘available’ for signaling. Conversely, the MDA5 CARDs are unhindered in the absence of viral RNA.[25] As a safeguard for RLR activation, the exposed RIG-I and MDA5 CARDs can undergo post-translational modifications (e.g. ubiquitination, phosphorylation) that either positively or negatively regulate downstream signaling.
RIG-I antiviral signaling
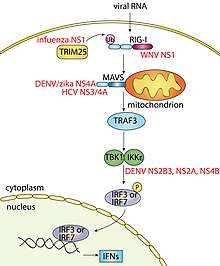
In the activated state the exposed RIG-I CARD domains interact with the CARD domains of MAVS (mitochondrial antiviral signaling protein, also known as IPS-1, VISA or Cardif) which sits on the outer surface of the mitochondria. This binding event is essential to signaling as it causes MAVS to form large functional aggregates in which TRAF3 (TNF receptor-associated factor 3) and subsequently the IKKε/TBK1 (I-kappa-B kinase-epsilon/TANK-binding kinase 1) complex are recruited. The IKKε/TBK1 complex leads to the activation of the transcription factors interferon regulatory factor 3 (IRF3) and IRF7 which induce type I (including IFNα and IFNβ) and type III interferons (IFN). The type I IFNs bind type I IFN receptors on the surface of the cell that produced them, and also other cell types that express the receptor, to activate JAK-STAT (Janus kinase/signal transducers and activators of transcription) signaling. This leads to the induction of hundreds of interferon stimulated genes (ISGs) that amplify the IFN response. Overall this causes the death of infected cells, the protection of surrounding cells and the activation of the antigen-specific antiviral immune response. Collectively this coordinated antiviral immune response controls the viral infection.
Regulation
As prolonged IFN production is linked to human disease RLR signaling must be tightly regulated. One of various ways that this is achieved is by post-translationally modifying, or tagging, host RLR signaling proteins with phosphate (known as phosphorylation) or ubiquitin (known as ubiquitination). These tags can also be removed, which adds an additional regulatory layer to RLR signaling. These post-translational modifications, and their removal, are prevalent in RLR signaling and even regulate the RIG-I receptor itself. Most famously the RIG-I CARD domain is phosphorylated by protein kinase C-α (PKC-α) and PKC-β in the resting state to negatively regulate signaling.[26][27][28] Upon viral infection RIG-I is dephosphorylated by PP1α and PP1γ,[29] permitting the ubiquitination of the RIG-I CARD domain by the E3 ligase TRIM25 to activate the RLR-mediated antiviral immune response.[30] Given post-translational modifications are so pertinent to the activation of RLR signaling, it is not surprising that they are directly, or indirectly, targeted by viruses such as influenza A[31] and measles,[32] respectively, to suppress signaling.
Viral hijacking of RLR signaling
Viruses have evolved ways to subvert RLR signaling to enhance their survival. For example, influenza A virus and West Nile virus (WNV) use their NS1 (nonstructural protein 1) proteins to block RIG-I ubiquitination by TRIM25, or cause RIG-I degradation, respectively, which in turn inhibits IFN production.[31][33] This outcome is also achieved by the hepatitis C (HCV) NS3/4A protein by cleaving a part of MAVS,[34] and the foot-and-mouth disease virus (FMDV) leader protease (Lpro) which cleaves LGP2.[35] Likewise, dengue virus (DENV) uses its NS2B3, NS2A and NS4B proteins to bind IKKε and prevent IRF3 phosphorylation[36][37] and its NS4A protein, as per the zika virus, to bind MAVS to block RLR receptor binding.[38][39] Another prominent example is that of the paramyxovirus V proteins, which directly bind various RLR or downstream signaling proteins including MDA5, LGP2, and STAT,[40][41][42] or proteins such as PP1α and PP1γ[32] that negatively regulate RLR signaling.
See also
- NOD-like receptor
- Toll-like receptor
References
- Mahla RS, Reddy MC, Prasad DV, Kumar H (September 2013). "Sweeten PAMPs: Role of Sugar Complexed PAMPs in Innate Immunity and Vaccine Biology". Frontiers in Immunology. 4: 248. doi:10.3389/fimmu.2013.00248. PMC 3759294. PMID 24032031.
- Stefan Offermanns; Walter Rosenthal (2008-08-14). Encyclopedia of Molecular Pharmacology, Volume 1. Springer. ISBN 9783540389163. Retrieved 30 August 2011.
The other two families of PRRs, the NOD-like receptors (NLRs) and the RIG-like helicases (RLHs) are soluble receptors present in the cytosol and act as sensors to detect a variety of viral and bacterial products.
- Kato, Hiroki; Takeuchi, Osamu; Mikamo-Satoh, Eriko; Hirai, Reiko; Kawai, Tomoji; Matsushita, Kazufumi; Hiiragi, Akane; Dermody, Terence S.; Fujita, Takashi (2008-06-30). "Length-dependent recognition of double-stranded ribonucleic acids by retinoic acid–inducible gene-I and melanoma differentiation–associated gene 5". The Journal of Experimental Medicine. 205 (7): 1601–1610. doi:10.1084/jem.20080091. ISSN 0022-1007. PMC 2442638. PMID 18591409.
- Saito, Takeshi; Owen, David M.; Jiang, Fuguo; Marcotrigiano, Joseph; Gale, Michael (2008-06-11). "Innate immunity induced by composition-dependent RIG-I recognition of hepatitis C virus RNA". Nature. 454 (7203): 523–527. doi:10.1038/nature07106. ISSN 0028-0836. PMC 2856441. PMID 18548002.
- Baum, A.; Sachidanandam, R.; Garcia-Sastre, A. (2010-08-30). "Preference of RIG-I for short viral RNA molecules in infected cells revealed by next-generation sequencing". Proceedings of the National Academy of Sciences. 107 (37): 16303–16308. doi:10.1073/pnas.1005077107. ISSN 0027-8424. PMC 2941304. PMID 20805493.
- Gitlin, L.; Barchet, W.; Gilfillan, S.; Cella, M.; Beutler, B.; Flavell, R. A.; Diamond, M. S.; Colonna, M. (2006-05-19). "Essential role of mda-5 in type I IFN responses to polyriboinosinic:polyribocytidylic acid and encephalomyocarditis picornavirus". Proceedings of the National Academy of Sciences. 103 (22): 8459–8464. doi:10.1073/pnas.0603082103. ISSN 0027-8424. PMC 1464000. PMID 16714379.
- Hornung, V.; Ellegast, J.; Kim, S.; Brzozka, K.; Jung, A.; Kato, H.; Poeck, H.; Akira, S.; Conzelmann, K.-K. (2006-11-10). "5'-Triphosphate RNA Is the Ligand for RIG-I". Science. 314 (5801): 994–997. doi:10.1126/science.1132505. ISSN 0036-8075. PMID 17038590.
- Wang, Haifeng; Ryu, Wang-Shick (2010-07-15). "Hepatitis B Virus Polymerase Blocks Pattern Recognition Receptor Signaling via Interaction with DDX3: Implications for Immune Evasion". PLoS Pathogens. 6 (7): e1000986. doi:10.1371/journal.ppat.1000986. ISSN 1553-7374. PMC 2904777. PMID 20657822.
- Schlee, Martin; Roth, Andreas; Hornung, Veit; Hagmann, Cristina Amparo; Wimmenauer, Vera; Barchet, Winfried; Coch, Christoph; Janke, Markus; Mihailovic, Aleksandra (2009). "Recognition of 5′ Triphosphate by RIG-I Helicase Requires Short Blunt Double-Stranded RNA as Contained in Panhandle of Negative-Strand Virus". Immunity. 31 (1): 25–34. doi:10.1016/j.immuni.2009.05.008. ISSN 1074-7613. PMC 2824854. PMID 19576794.
- Feng, Qian; Hato, Stanleyson V.; Langereis, Martijn A.; Zoll, Jan; Virgen-Slane, Richard; Peisley, Alys; Hur, Sun; Semler, Bert L.; van Rij, Ronald P. (2012). "MDA5 Detects the Double-Stranded RNA Replicative Form in Picornavirus-Infected Cells". Cell Reports. 2 (5): 1187–1196. doi:10.1016/j.celrep.2012.10.005. ISSN 2211-1247. PMID 23142662.
- Kato, Hiroki; Takeuchi, Osamu; Sato, Shintaro; Yoneyama, Mitsutoshi; Yamamoto, Masahiro; Matsui, Kosuke; Uematsu, Satoshi; Jung, Andreas; Kawai, Taro (2006-04-09). "Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses". Nature. 441 (7089): 101–105. doi:10.1038/nature04734. ISSN 0028-0836. PMID 16625202.
- Li, Xiaojun; Ranjith-Kumar, C. T.; Brooks, Monica T.; Dharmaiah, S.; Herr, Andrew B.; Kao, Cheng; Li, Pingwei (2009-03-11). "The RIG-I-like Receptor LGP2 Recognizes the Termini of Double-stranded RNA". Journal of Biological Chemistry. 284 (20): 13881–13891. doi:10.1074/jbc.m900818200. ISSN 0021-9258. PMC 2679488. PMID 19278996.
- Murali, Ayaluru; Li, Xiaojun; Ranjith-Kumar, C. T.; Bhardwaj, Kanchan; Holzenburg, Andreas; Li, Pingwei; Kao, C. Cheng (2008-04-14). "Structure and Function of LGP2, a DEX(D/H) Helicase That Regulates the Innate Immunity Response". Journal of Biological Chemistry. 283 (23): 15825–15833. doi:10.1074/jbc.m800542200. ISSN 0021-9258. PMC 2414291. PMID 18411269.
- Bruns, Annie M.; Leser, George P.; Lamb, Robert A.; Horvath, Curt M. (2014). "The Innate Immune Sensor LGP2 Activates Antiviral Signaling by Regulating MDA5-RNA Interaction and Filament Assembly". Molecular Cell. 55 (5): 771–781. doi:10.1016/j.molcel.2014.07.003. ISSN 1097-2765. PMC 4156907. PMID 25127512.
- Satoh, T.; Kato, H.; Kumagai, Y.; Yoneyama, M.; Sato, S.; Matsushita, K.; Tsujimura, T.; Fujita, T.; Akira, S. (2010-01-08). "LGP2 is a positive regulator of RIG-I- and MDA5-mediated antiviral responses". Proceedings of the National Academy of Sciences. 107 (4): 1512–1517. doi:10.1073/pnas.0912986107. ISSN 0027-8424. PMC 2824407. PMID 20080593.
- Takahasi, Kiyohiro; Kumeta, Hiroyuki; Tsuduki, Natsuko; Narita, Ryo; Shigemoto, Taeko; Hirai, Reiko; Yoneyama, Mitsutoshi; Horiuchi, Masataka; Ogura, Kenji (2009-04-20). "Solution Structures of Cytosolic RNA Sensor MDA5 and LGP2 C-terminal Domains". Journal of Biological Chemistry. 284 (26): 17465–17474. doi:10.1074/jbc.m109.007179. ISSN 0021-9258. PMC 2719387. PMID 19380577.
- Bruns, Annie M.; Leser, George P.; Lamb, Robert A.; Horvath, Curt M. (2014). "The Innate Immune Sensor LGP2 Activates Antiviral Signaling by Regulating MDA5-RNA Interaction and Filament Assembly". Molecular Cell. 55 (5): 771–781. doi:10.1016/j.molcel.2014.07.003. ISSN 1097-2765. PMC 4156907. PMID 25127512.
- Childs, Kay S.; Randall, Richard E.; Goodbourn, Stephen (2013-05-09). "LGP2 Plays a Critical Role in Sensitizing mda-5 to Activation by Double-Stranded RNA". PLoS ONE. 8 (5): e64202. doi:10.1371/journal.pone.0064202. ISSN 1932-6203. PMID 23671710.
- Komuro, A.; Horvath, C. M. (2006-10-04). "RNA- and Virus-Independent Inhibition of Antiviral Signaling by RNA Helicase LGP2". Journal of Virology. 80 (24): 12332–12342. doi:10.1128/jvi.01325-06. ISSN 0022-538X.
- Parisien, Jean‐Patrick; Lenoir, Jessica J; Mandhana, Roli; Rodriguez, Kenny R; Qian, Kenin; Bruns, Annie M; Horvath, Curt M (2018-04-16). "RNA sensor LGP2 inhibits TRAF ubiquitin ligase to negatively regulate innate immune signaling". EMBO Reports. 19 (6): e45176. doi:10.15252/embr.201745176. ISSN 1469-221X. PMC 5989757. PMID 29661858.
- Saito, T.; Hirai, R.; Loo, Y.-M.; Owen, D.; Johnson, C. L.; Sinha, S. C.; Akira, S.; Fujita, T.; Gale, M. (2006-12-26). "Regulation of innate antiviral defenses through a shared repressor domain in RIG-I and LGP2". Proceedings of the National Academy of Sciences. 104 (2): 582–587. doi:10.1073/pnas.0606699104. ISSN 0027-8424. PMC 1766428. PMID 17190814.
- Satoh, T.; Kato, H.; Kumagai, Y.; Yoneyama, M.; Sato, S.; Matsushita, K.; Tsujimura, T.; Fujita, T.; Akira, S. (2010-01-08). "LGP2 is a positive regulator of RIG-I- and MDA5-mediated antiviral responses". Proceedings of the National Academy of Sciences. 107 (4): 1512–1517. doi:10.1073/pnas.0912986107. ISSN 0027-8424. PMC 2824407. PMID 20080593.
- Uchikawa, Emiko; Lethier, Mathilde; Malet, Hélène; Brunel, Joanna; Gerlier, Denis; Cusack, Stephen (2016). "Structural Analysis of dsRNA Binding to Anti-viral Pattern Recognition Receptors LGP2 and MDA5". Molecular Cell. 62 (4): 586–602. doi:10.1016/j.molcel.2016.04.021. ISSN 1097-2765. PMID 27203181.
- Luo, Dahai; Ding, Steve C.; Vela, Adriana; Kohlway, Andrew; Lindenbach, Brett D.; Pyle, Anna Marie (2011). "Structural Insights into RNA Recognition by RIG-I". Cell. 147 (2): 409–422. doi:10.1016/j.cell.2011.09.023. ISSN 0092-8674. PMID 22000018.
- Berke, Ian C; Modis, Yorgo (2012-02-07). "MDA5 cooperatively forms dimers and ATP-sensitive filaments upon binding double-stranded RNA". The EMBO Journal. 31 (7): 1714–1726. doi:10.1038/emboj.2012.19. ISSN 0261-4189. PMC 3321199. PMID 22314235.
- Gack, M. U.; Nistal-Villan, E.; Inn, K.-S.; Garcia-Sastre, A.; Jung, J. U. (2010-01-13). "Phosphorylation-Mediated Negative Regulation of RIG-I Antiviral Activity". Journal of Virology. 84 (7): 3220–3229. doi:10.1128/jvi.02241-09. ISSN 0022-538X. PMID 20071582.
- Maharaj, N. P.; Wies, E.; Stoll, A.; Gack, M. U. (2011-11-23). "Conventional Protein Kinase C- (PKC- ) and PKC- Negatively Regulate RIG-I Antiviral Signal Transduction". Journal of Virology. 86 (3): 1358–1371. doi:10.1128/jvi.06543-11. ISSN 0022-538X.
- Nistal-Villán, Estanislao; Gack, Michaela U.; Martínez-Delgado, Gustavo; Maharaj, Natalya P.; Inn, Kyung-Soo; Yang, Heyi; Wang, Rong; Aggarwal, Aneel K.; Jung, Jae U. (2010-04-20). "Negative Role of RIG-I Serine 8 Phosphorylation in the Regulation of Interferon-β Production". Journal of Biological Chemistry. 285 (26): 20252–20261. doi:10.1074/jbc.m109.089912. ISSN 0021-9258.
- Wies, Effi; Wang, May K.; Maharaj, Natalya P.; Chen, Kan; Zhou, Shenghua; Finberg, Robert W.; Gack, Michaela U. (2013). "Dephosphorylation of the RNA Sensors RIG-I and MDA5 by the Phosphatase PP1 Is Essential for Innate Immune Signaling". Immunity. 38 (3): 437–449. doi:10.1016/j.immuni.2012.11.018. ISSN 1074-7613.
- Gack, Michaela U.; Shin, Young C.; Joo, Chul-Hyun; Urano, Tomohiko; Liang, Chengyu; Sun, Lijun; Takeuchi, Osamu; Akira, Shizuo; Chen, Zhijian (2007-03-28). "TRIM25 RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated antiviral activity". Nature. 446 (7138): 916–920. doi:10.1038/nature05732. ISSN 0028-0836.
- Gack, Michaela Ulrike; Albrecht, Randy Allen; Urano, Tomohiko; Inn, Kyung-Soo; Huang, I-Chueh; Carnero, Elena; Farzan, Michael; Inoue, Satoshi; Jung, Jae Ung (2005). "Influenza A Virus NS1 Targets the Ubiquitin Ligase TRIM25 to Evade Recognition by the Host Viral RNA Sensor RIG-I". Cell Host & Microbe. 5 (5): 439–449. doi:10.1016/j.chom.2009.04.006. ISSN 1931-3128.
- Davis, Meredith E.; Wang, May K.; Rennick, Linda J.; Full, Florian; Gableske, Sebastian; Mesman, Annelies W.; Gringhuis, Sonja I.; Geijtenbeek, Teunis B.H.; Duprex, W. Paul (2014). "Antagonism of the Phosphatase PP1 by the Measles Virus V Protein Is Required for Innate Immune Escape of MDA5". Cell Host & Microbe. 16 (1): 19–30. doi:10.1016/j.chom.2014.06.007. ISSN 1931-3128.
- Zhang, Hong-Lei; Ye, Han-Qing; Liu, Si-Qing; Deng, Cheng-Lin; Li, Xiao-Dan; Shi, Pei-Yong; Zhang, Bo (2017-06-28). "West Nile Virus NS1 Antagonizes Interferon Beta Production by Targeting RIG-I and MDA5". Journal of Virology. 91 (18). doi:10.1128/jvi.02396-16. ISSN 0022-538X.
- Li, X.-D.; Sun, L.; Seth, R. B.; Pineda, G.; Chen, Z. J. (2005-11-21). "Hepatitis C virus protease NS3/4A cleaves mitochondrial antiviral signaling protein off the mitochondria to evade innate immunity". Proceedings of the National Academy of Sciences. 102 (49): 17717–17722. doi:10.1073/pnas.0508531102. ISSN 0027-8424.
- Rodríguez Pulido, Miguel; Sánchez-Aparicio, María Teresa; Martínez-Salas, Encarnación; García-Sastre, Adolfo; Sobrino, Francisco; Sáiz, Margarita (2018-06-29). "Innate immune sensor LGP2 is cleaved by the Leader protease of foot-and-mouth disease virus". PLOS Pathogens. 14 (6): e1007135. doi:10.1371/journal.ppat.1007135. ISSN 1553-7374.
- Angleró-Rodríguez, Yesseinia I.; Pantoja, Petraleigh; Sariol, Carlos A. (2013-10-30). "Dengue Virus Subverts the Interferon Induction Pathway via NS2B/3 Protease-IκB Kinase ε Interaction". Clinical and Vaccine Immunology. 21 (1): 29–38. doi:10.1128/cvi.00500-13. ISSN 1556-6811.
- Dalrymple, Nadine A.; Cimica, Velasco; Mackow, Erich R. (2015-05-12). "Dengue Virus NS Proteins Inhibit RIG-I/MAVS Signaling by Blocking TBK1/IRF3 Phosphorylation: Dengue Virus Serotype 1 NS4A Is a Unique Interferon-Regulating Virulence Determinant". mBio. 6 (3). doi:10.1128/mbio.00553-15. ISSN 2150-7511.
- He, Zhenjian; Zhu, Xun; Wen, Weitao; Yuan, Jie; Hu, Yiwen; Chen, Jiahui; An, Shu; Dong, Xinhuai; Lin, Cuiji (2016-06-01). "Dengue Virus Subverts Host Innate Immunity by Targeting Adaptor Protein MAVS". Journal of Virology. 90 (16): 7219–7230. doi:10.1128/jvi.00221-16. ISSN 0022-538X. PMC 4984625. PMID 27252539.
- Ma, Jinzhu; Ketkar, Harshada; Geng, Tingting; Lo, Emily; Wang, Leilei; Xi, Juemin; Sun, Qiangming; Zhu, Zhanbo; Cui, Yudong (2018-06-25). "Zika Virus Non-structural Protein 4A Blocks the RLR-MAVS Signaling". Frontiers in Microbiology. 9. doi:10.3389/fmicb.2018.01350. ISSN 1664-302X.
- Andrejeva, J.; Childs, K. S.; Young, D. F.; Carlos, T. S.; Stock, N.; Goodbourn, S.; Randall, R. E. (2004-11-24). "The V proteins of paramyxoviruses bind the IFN-inducible RNA helicase, mda-5, and inhibit its activation of the IFN- promoter". Proceedings of the National Academy of Sciences. 101 (49): 17264–17269. doi:10.1073/pnas.0407639101. ISSN 0027-8424. PMC 535396. PMID 15563593.
- Childs, Kay; Stock, Nicola; Ross, Craig; Andrejeva, Jelena; Hilton, Louise; Skinner, Michael; Randall, Richard; Goodbourn, Stephen (2007). "mda-5, but not RIG-I, is a common target for paramyxovirus V proteins". Virology. 359 (1): 190–200. doi:10.1016/j.virol.2006.09.023. ISSN 0042-6822. PMID 17049367.
- Rodriguez, Kenny R.; Horvath, Curt M. (2014-05-14). "Paramyxovirus V Protein Interaction with the Antiviral Sensor LGP2 Disrupts MDA5 Signaling Enhancement but Is Not Relevant to LGP2-Mediated RLR Signaling Inhibition". Journal of Virology. 88 (14): 8180–8188. doi:10.1128/jvi.00737-14. ISSN 0022-538X. PMC 4097766. PMID 24829334.
External links
- PTHR14074: HELICASE WITH DEATH DOMAIN-RELATED (filter for human)