Pulmonary surfactant
Pulmonary surfactant is a surface-active lipoprotein complex (phospholipoprotein) formed by type II alveolar cells. The proteins and lipids that make up the surfactant have both hydrophilic and hydrophobic regions. By adsorbing to the air-water interface of alveoli, with hydrophilic head groups in the water and the hydrophobic tails facing towards the air, the main lipid component of surfactant, dipalmitoylphosphatidylcholine (DPPC), reduces surface tension.
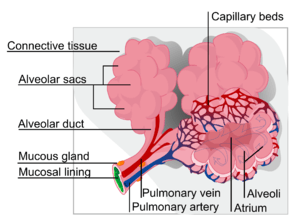
As a medication, pulmonary surfactant is on the WHO Model List of Essential Medicines, the most important medications needed in a basic health system.[1]
Function
- To increase pulmonary compliance.
- To prevent atelectasis (collapse of the lung) at the end of expiration.
- To facilitate recruitment of collapsed airways.
Alveoli can be compared to gas in water, as the alveoli are wet and surround a central air space. The surface tension acts at the air-water interface and tends to make the bubble smaller (by decreasing the surface area of the interface). The gas pressure (P) needed to keep equilibrium between the collapsing force of surface tension (γ) and the expanding force of gas in an alveolus of radius r is expressed by the law of Laplace:

Compliance
Compliance is the ability of lungs and thorax to expand. Lung compliance is defined as the volume change per unit of pressure change across the lung. Measurements of lung volume obtained during the controlled inflation/deflation of a normal lung show that the volumes obtained during deflation exceed those during inflation, at a given pressure. This difference in inflation and deflation volumes at a given pressure is called hysteresis and is due to the air-water surface tension that occurs at the beginning of inflation. However, surfactant decreases the alveolar surface tension, as seen in cases of premature infants suffering from infant respiratory distress syndrome. The normal surface tension for water is 70 dyn/cm (70 mN/m) and in the lungs it is 25 dyn/cm (25 mN/m); however, at the end of the expiration, compressed surfactant phospholipid molecules decrease the surface tension to very low, near-zero levels. Pulmonary surfactant thus greatly reduces surface tension, increasing compliance allowing the lung to inflate much more easily, thereby reducing the work of breathing. It reduces the pressure difference needed to allow the lung to inflate. The lung's compliance decreases and ventilation decreases when lung tissue becomes diseased and fibrotic.[2]
Alveolar size regulation
As the alveoli increase in size, the surfactant becomes more spread out over the surface of the liquid. This increases surface tension effectively slowing the rate of expansion of the alveoli. This also helps all alveoli in the lungs expand at the same rate, as one that expands more quickly will experience a large rise in surface tension slowing its rate of expansion. It also means the rate of shrinking is more regular, as if one reduces in size more quickly the surface tension will reduce more, so other alveoli can contract more easily than it can. Surfactant reduces surface tension more readily when the alveoli are smaller because the surfactant is more concentrated.
Prevention of fluid accumulation and maintenance of dryness of airways
Surface tension draws fluid from capillaries to the alveolar spaces. Surfactant reduces fluid accumulation and keeps the airways dry by reducing surfacee tension.[3]
Innate immunity
Surfactant immune function is primarily attributed to two proteins: SP-A and SP-D. These proteins can bind to sugars on the surface of pathogens and thereby opsonize them for uptake by phagocytes. It also regulates inflammatory responses and interacts with the adaptive immune response. Surfactant degradation or inactivation may contribute to enhanced susceptibility to lung inflammation and infection.[4]
Composition
- ~40% dipalmitoylphosphatidylcholine (DPPC);[5]
- ~40% other phospholipids (PC);[5]
- ~10% surfactant proteins (SP-A, SP-B, SP-C and SP-D);[5]
- ~10% neutral lipids (Cholesterol);[5]
- Traces of other substances.
Lipids
DPPC
Dipalmitoylphosphatidylcholine (DPPC) is a phospholipid with two 16-carbon saturated chains and a phosphate group with quaternary amine group attached. The DPPC is the strongest surfactant molecule in the pulmonary surfactant mixture. It also has higher compaction capacity than the other phospholipids, because the apolar tail is less bent. Nevertheless, without the other substances of the pulmonary surfactant mixture, the DPPC's adsorption kinetics is very slow. This happens primarily because the phase transition temperature between gel to liquid crystal of pure DPPC is 41.5 °C, which is higher than the human body's temperature of 37 °C.[6]
Other phospholipids
Phosphatidylcholine molecules form ~85% of the lipid in surfactant and have saturated acyl chains. Phosphatidylglycerol (PG) forms about 11% of the lipids in the surfactant, it has unsaturated fatty acid chains that fluidize the lipid monolayer at the interface. Neutral lipids and cholesterol are also present. The components for these lipids diffuse from the blood into type II alveolar cells where they are assembled and packaged for secretion into secretory organelles called lamellar bodies.
Proteins
Proteins make up the remaining 10% of the surfactant. Half of this 10% is plasma proteins but the rest is formed by the apolipoproteins, surfactant proteins SP-A, SP-B, SP-C, and SP-D. The apolipoproteins are produced by the secretory pathway in type II cells. They undergo much post-translational modification, ending up in the lamellar bodies. These are concentric rings of lipid and protein, about 1 µm in diameter.
- SP-A and SP-D are collectins. They confer innate immunity as they have carbohydrate recognition domains that allow them to coat bacteria and viruses promoting phagocytosis by macrophages.[7] SP-A is also thought to be involved in a negative feedback mechanism to control the production of surfactant.
- SP-B and SP-C are hydrophobic membrane proteins that increase the rate that surfactant spreads over the surface. SP-B and SP-C are required for proper biophysical function of the lung. Humans and animals born with a congenital absence of the saposin family SP-B suffer from intractable respiratory failure whereas those born lacking SP-C tend to develop progressive interstitial pneumonitis.[8]
The SP proteins reduce the critical temperature of DPPC's phase transition to a value lower than 37 °C,[9] which improves its adsorption and interface spreading velocity.[10][11] The compression of the interface causes a phase change of the surfactant molecules to liquid-gel or even gel-solid. The fast adsorption velocity is necessary to maintain the integrity of the gas exchange region of the lungs.
Each SP protein has distinct functions, which act synergistically to keep an interface rich in DPPC during lung's expansion and contraction. Changes in the surfactant mixture composition alter the pressure and temperature conditions for phase changes and the phospholipids' crystal shape as well.[12] Only the liquid phase can freely spread on the surface to form a monolayer. Nevertheless, it has been observed that if a lung region is abruptly expanded the floating crystals crack like "icebergs". Then the SP proteins selectively attract more DPPC to the interface than other phospholipids or cholesterol, whose surfactant properties are worse than DPPC's. The SP also fastens the DPPC on the interface to prevent the DPPC from being squeezed out when the surface area decreases [11] This also reduces the interface compressibility.[13]
Artificial surfactants

There are a number of types of pulmonary surfactants available.
Synthetic pulmonary surfactants
- Colfosceril palmitate (Exosurf) - a mixture of DPPC with hexadecanol and tyloxapol added as spreading agents
- Pumactant (Artificial Lung Expanding Compound or ALEC) - a mixture of DPPC and PG
- KL-4 - composed of DPPC, palmitoyl-oleoyl phosphatidylglycerol, and palmitic acid, combined with a 21 amino acid synthetic peptide that mimics the structural characteristics of SP-B.
- Venticute - DPPC, PG, palmitic acid and recombinant SP-C
- Lucinactant - DPPC, POPG, and palmitic acid.
Animal derived surfactants
- Beractant
- (Alveofact) - extracted from cow lung lavage fluid
- (Survanta) - extracted from minced cow lung with additional DPPC, palmitic acid and tripalmitin
- (Beraksurf) -extracted from minced calf lung with additional DPPC, palmitic acid and tripalmitin
- Calfactant (Infasurf) - extracted from calf lung lavage fluid
- Poractant alfa (Curosurf) - extracted from material derived from minced pig lung
Surface tension magnitude inside the lung
Even though the surface tension can be greatly reduced by pulmonary surfactant, this effect will depend on the surfactant's concentration on the interface. The interface concentration has a saturation limit, which depends on temperature and mixture composition. Because during ventilation there is a variation of the lung surface area, the surfactant's interface concentration is not usually at the level of saturation. The surface increases during inspiration, which consequently opens space for new surfactant molecules to be recruited to the interface. Meanwhile, at the expiration the surface area decreases, the layer of surfactant is squeezed, bringing the surfactant molecules closer to each other and further decreasing the surface tension.
SP molecules contribute to increase the surfactant interface adsorption kinetics, when the concentration is below the saturation level. They also make weak bonds with the surfactant molecules at the interface and hold them longer there when the interface is compressed. Therefore, during ventilation, surface tension is usually lower than at equilibrium. Therefore, the surface tension varies according to the volume of air in the lungs, which protects them from atelectasis at low volumes and tissue damage at high volume levels.[10][12][13]
| Condition | Tension (mN/m) |
|---|---|
| Water at 25 °C | 70 |
| Pulmonary surfactant in equilibrium at 36 °C | 25 |
| Healthy lung at 100% of TLC | 30 |
| Healthy lung between 40 and 60% of TLC | 1~6 |
| Healthy lung below 40% of TLC | <1 |
Production and degradation
Surfactant production in humans begins in Type II cells during the alveolar sac stage of lung development. Lamellar bodies appear in the cytoplasm at about 20 weeks gestation. These lamellar bodies are secreted by exocytosis into the surface water layer lining the alveolar airspace, where the surfactant forms a meshwork of tubular myelin. Term infants are estimated to have an alveolar storage pool of approximately 100 mg/kg of surfactant, while preterm infants have an estimated 4–5 mg/kg at birth.
Club cells also produce a component of lung surfactant.[14]
Alveolar surfactant has a half life of 5 to 10 hours once secreted. It can be both broken down by macrophages and / or reabsorbed into the lamellar structures of type II pneumocytes. Up to 90% of surfactant DPPC (dipalmitoyl phosphatidylcholine) is recycled from the alveolar space back into the type II pneumocyte. This process is believed to occur through SP-A stimulating receptor mediated, clathrin dependent endocytosis.[15] The other 10% is taken up by alveolar macrophages and digested.
Diseases
- Infant respiratory distress syndrome (IRDS) is caused by lack of surfactant, commonly suffered by premature babies born before 28–32 weeks of gestation.
- Congenital surfactant deficiency
- Pulmonary alveolar proteinosis
- Surfactant metabolism dysfunction
History
In late 1920s von Neergaard[16] identified the function of the pulmonary surfactant in increasing the compliance of the lungs by reducing surface tension. However the significance of his discovery was not understood by the scientific and medical community at that time. He also realized the importance of having low surface tension in lungs of newborn infants. Later, in the middle of the 1950s, Pattle and Clements rediscovered the importance of surfactant and low surface tension in the lungs. At the end of that decade it was discovered that the lack of surfactant caused infant respiratory distress syndrome (IRDS).[12]
References
- "19th WHO Model List of Essential Medicines (April 2015)" (PDF). WHO. April 2015. Retrieved May 10, 2015.
- "Alveoli and the Breathing Process". Retrieved 2013-10-30.
- West, John B. (1994). Respiratory physiology-- the essentials. Baltimore: Williams & Wilkins. ISBN 0-683-08937-4.
- Wright, Jo Rae (2004). "Host Defense Functions of Pulmonary Surfactant". Biology of the Neonate. 85 (4): 326–32. doi:10.1159/000078172. PMID 15211087.
- Nkadi, Paul O.; Merritt, T. Allen; Pillers, De-Ann M. (2009). "An overview of pulmonary surfactant in the neonate: Genetics, metabolism, and the role of surfactant in health and disease". Molecular Genetics and Metabolism. 97 (2): 95–101. doi:10.1016/j.ymgme.2009.01.015. ISSN 1096-7192. PMC 2880575.
- Albon, Norman (1978). "Nature of the gel to liquid crystal transition of synthetic phosphatidylcholines". Proceedings of the National Academy of Sciences of the United States of America. 75 (5): 2258–2260. doi:10.1073/pnas.75.5.2258. PMC 392531. PMID 276866.
- Haagsman HP, Diemel RV (May 2001). "Surfactant-associated proteins: functions and structural variation". Comparative Biochemistry and Physiology. Part A, Molecular & Integrative Physiology. 129 (1): 91–108. doi:10.1016/s1095-6433(01)00308-7. PMID 11369536.
- Weaver TE, Conkright JJ (2001). "Function of surfactant proteins B and C". Annual Review of Physiology. 63: 555–78. doi:10.1146/annurev.physiol.63.1.555. PMID 11181967.
- Hills, B. A. (1999). "An alternative view of the role(s) of surfactant and the alveolar model". Journal of Applied Physiology. 87 (5): 1567–83. doi:10.1152/jappl.1999.87.5.1567. PMID 10562593.
- Schurch, S.; Lee, Martin; Gehr, Peter; Qanbar, R; Schürch, S (1992). "Pulmonary surfactant: Surface properties and function of alveolar and airway surfactant". Pure and Applied Chemistry. 64 (11): 209–20. doi:10.1351/pac199264111745.
- Possmayer, Fred; Nag, Kaushik; Rodriguez, Karina; Qanbar, Riad; Schürch, Samuel (2001). "Surface activity in vitro: Role of surfactant proteins". Comparative Biochemistry and Physiology A. 129: 209–20. doi:10.1016/S1095-6433(01)00317-8. PMID 11369545.
- Veldhuizen, Ruud; Nag, Kaushik; Orgeig, Sandra; Possmayer, Fred (1998). "The role of lipids in pulmonary surfactant". Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease. 1408 (2–3): 90–108. doi:10.1016/S0925-4439(98)00061-1. PMID 9813256.
- Schürch, Samuel; Bachofen, Hans; Possmayer, Fred (2001). "Surface activity in situ, in vivo, and in the captive bubble surfactometer". Comparative Biochemistry and Physiology A. 129 (1): 195–207. doi:10.1016/S1095-6433(01)00316-6. PMID 11369544.
- Young, Barbara (2014). Wheater's functional histology : a text and colour atlas. O'Dowd, Geraldine,, Woodford, Phillip, (Sixth ed.). Philadelphia, PA. pp. Ch 12. ISBN 0702047473. OCLC 861650889.CS1 maint: extra punctuation (link)
- Crowther, J. E.; Schlesinger, L. S. (2005). "Endocytic pathway for surfactant protein a in human macrophages: Binding, clathrin-mediated uptake, and trafficking through the endolysosomal pathway". AJP: Lung Cellular and Molecular Physiology. 290 (2): L334–42. doi:10.1152/ajplung.00267.2005. PMID 16169899.
- Neergaard, K. (1929). "Neue Auffassungen über einen Grundbegriff der Atemmechanik" [New views on a fundamental concept of respiratory mechanics]. Zeitschrift für Die Gesamte Experimentelle Medizin (in German). 66 (1): 373–94. doi:10.1007/bf02621963.