Physarum polycephalum
Physarum polycephalum, an acellular slime mold or myxomycete, is a protist with diverse cellular forms and broad geographic distribution. The “acellular” moniker derives from the plasmodial stage of the life cycle: the plasmodium is a bright yellow macroscopic multinucleate syncytium. This stage of the life cycle, along with its preference for damp shady habitats, likely contributed to the original mischaracterization of the organism as a fungus. P. polycephalum is used as a model organism for research into motility, cellular differentiation, chemotaxis, cellular compatibility, and the cell cycle.
| Physarum polycephalum | |
|---|---|
 | |
| Physarum polycephalum plasmodium forming over tree chunk. | |
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Order: | Physarales |
| Family: | Physaraceae |
| Genus: | |
| Species: | P. polycephalum |
| Binomial name | |
| Physarum polycephalum | |
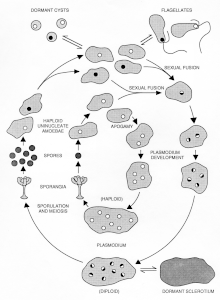
Life cycle and characteristics
The two vegetative cell types, amoebae and plasmodia, differ markedly in morphology, physiology and behavior. Amoebae are microorganisms, typically haploid, that live primarily in the soil, where they phagocytose bacteria. In the laboratory amoebae are grown on lawns of live or dead Escherichia coli on nutrient agar plates, where they can multiply indefinitely. Axenic culture of amoebae was achieved through selection of mutants capable of axenic growth.[1] Under conditions of starvation or desiccation, the amoebae differentiate reversibly into dormant spores with cell walls. When immersed in water, amoebae differentiate reversibly into flagellated cells, which involves a major reorganization of the cytoskeleton.[2]
The plasmodium is typically diploid and propagates via growth and nuclear division without cytokinesis, resulting in the macroscopic multinucleate syncytium. While nutrients are available, the plasmodium can grow to a foot or more in diameter. Like amoebae, the plasmodium can consume whole microbes, but also readily grows axenically in liquid cultures, nutrient agar plates and on nutrient-moistened surfaces. When nutrients are provided uniformly, the nuclei in the plasmodium divide synchronously, accounting for the interest in using P. polycephalum as a model organism to study the cell cycle, or more specifically the nuclear division cycle. When the plasmodium is starved, it has two alternative developmental pathways. In the dark, the plasmodium typically differentiates reversibly into a dormant “sclerotium” (the same term is used for dormant forms of fungal mycelia.[3] , but the myxomycete sclerotium is a very different structure). When exposed to light, the starving plasmodium differentiates irreversibly into sporangia that are distinguished from other Physarum species by their multiple heads (hence polycephalum). Meiosis occurs during spore development, resulting in haploid dormant spores. Upon exposure to moist nutrient conditions, the spores develop into amoebae, or, in aqueous suspension, into flagellates. The life cycle is completed when haploid amoebae of different mating types fuse to form a diploid zygote that then develops by growth and nuclear division in the absence of cytokinesis into the multinucleate plasmodium.[4]
In laboratory strains carrying a mutation at the matA mating type locus, the differentiation of P. polycephalum plasmodia can occur without the fusion of amoebae, resulting in haploid plasmodia that are morphologically indistinguishable from the more typical diploid form.[5] This enables facile genetic analysis of plasmodial traits that would otherwise require backcrossing to achieve homozygosity for analysis of recessive mutations in diploids. Sporangia from haploid plasmodia generate spores with low fertility, and it is assumed that viable spores develop from meiosis of rare diploid nuclei in the otherwise haploid P. polycephalum plasmodia. Apogamic development can also occur in nature in various species of myxomycetes.[6] In the figure of the P. polycephalum life cycle, the typical haploid-diploid sexual cycle is depicted in the outer circuit and the apogamic cycle in the inner circuit. Note that an apogamic amoeba retains its matA1 mating type speificity and can still fuse sexually with an amoeba of a different mating type to form a diploid heterozygous plasmodium—another characteristic that facilitates genetic analysis.
As the life cycle diagram indicates, amoebae and plasmodia differ markedly in their developmental potential. A remarkable further difference is the mechanism of mitosis. Amoebae exhibit “open mitosis” during which the nuclear membrane breaks down, as is typical of animal cells, before reassembling after telophase. Plasmodia exhibit “closed mitosis” during which the nuclear membrane remains intact. This presumably prevents nuclear fusion from occurring during mitosis in the multinucleate syncytium. In support of this inference, mutant amoebae defective in cytokinesis develop into multinucleate cells, and nuclear fusions during mitosis are common in these mutants.[7]
Streaming behavior
The movement of P. polycephalum is termed shuttle streaming. Shuttle streaming is characterized by the rhythmic back-and-forth flow of the protoplasm; the time interval is approximately two minutes. The forces of the streaming vary for each type of microplasmodium:
- The force in amoeboid microplasmodia is generated by contraction and relaxation of a membranous layer probably consisting of actin (a type of filament associated with contraction). The filament layer creates a pressure gradient, over which the protoplasm flows within the limits of the cell periphery.
- The force behind streaming in the dumbbell-shaped microplasmodia is generated by volume changes in both the periphery of the cell and in the invagination system of the cell membrane.
Situational behavior
Physarum polycephalum has been shown to exhibit characteristics similar to those seen in single-celled creatures and eusocial insects. For example, a team of Japanese and Hungarian researchers have shown P. polycephalum can solve the shortest path problem. When grown in a maze with oatmeal at two spots, P. polycephalum retracts from everywhere in the maze, except the shortest route connecting the two food sources.[8]
When presented with more than two food sources, P. polycephalum apparently solves a more complicated transportation problem. With more than two sources, the amoeba also produces efficient networks.[9] In a 2010 paper, oatflakes were dispersed to represent Tokyo and 36 surrounding towns.[10][11] P. polycephalum created a network similar to the existing train system, and "with comparable efficiency, fault tolerance, and cost". Similar results have been shown based on road networks in the United Kingdom[12] and the Iberian peninsula (i.e., Spain and Portugal).[13] P. polycephalum not only can solve these computational problems but also exhibits some form of memory. By repeatedly making the test environment of a specimen of P. polycephalum cold and dry for 60 minute intervals, Hokkaido University biophysicists discovered that the slime mold appears to anticipate the pattern by reacting to the conditions when they did not repeat the conditions for the next interval. Upon repeating the conditions, it would react to expect the 60 minute intervals, as well as testing with 30 and 90 minute intervals.[14][15]
P. polycephalum has also been shown to dynamically re-allocate to apparently maintain constant levels of different nutrients simultaneously.[16][17] In one particular instance, a specimen placed at the center of a Petri dish spatially re-allocated over combinations of food sources that each had different protein-carbohydrate ratios. After 60 hours, the slime mold area over each food source was measured. For each specimen, the results were consistent with the hypothesis that the amoeba would balance total protein and carbohydrate intake to reach particular levels that were invariant to the actual ratios presented to the slime mold.
As the slime mold does not have any nervous system that could explain these intelligent behaviours, there has been considerable interdisciplinary interest in understanding the rules that govern its behaviour. Scientists are trying to model the slime mold using a number of simple, distributed rules. For example, P. polycephalum has been modeled as a set of differential equations inspired by electrical networks. This model can be shown to be able to compute shortest paths.[18] A very similar model can be shown to solve the Steiner tree problem.[19] However, currently these models do not make sense biologically, as they for example assume energy conservation inside the slime mold.[lower-alpha 1] To build more realistic models, more data about the slime mold's network construction needs to be gathered. To this end, researchers are analysing the network structure of lab-grown P. polycephalum.[20]
In a book[21] and several preprints that have not been scientifically peer reviewed,[22][23] it has been claimed that because plasmodia appear to react in a consistent way to stimuli, they are the "ideal substrate for future and emerging bio-computing devices".[23] An outline has been presented showing how it may be possible to precisely point, steer and cleave plasmodium using light and food sources,[23] especially Valerian root.[24] Moreover, it has been reported that plasmodia can be made to form logic gates,[22] enabling the construction of biological computers. In particular, plasmodia placed at entrances to special geometrically shaped mazes would emerge at exits of the maze that were consistent with truth tables for certain primitive logic connectives. However, as these constructions are based on theoretical models of the slime mold, in practice these results do not scale to allow for actual computation. When the primitive logic gates are connected to form more complex functions, the plasmodium ceased to produce results consistent with the expected truth tables.
Even though complex computations using Physarum as a substrate are currently not possible, researchers have successfully used the organism's reaction to its environment in a USB sensor[25] and to control a robot.[26]
Footnotes
- Organisms consume food, so energy cannot be conserved.
References
- McCullough, Claire (1978). "Genetic Factors Determining the Growth of Physarum polycephalum Amoebae in Axenic Medium" (PDF). Journal of General Microbiology. 106 (2): 297–306. doi:10.1099/00221287-106-2-297 – via MicrobiologyResearch.org.
- Wright, Michel (1988). "Microtubule cytoskeleton and morphogenesis in the amoebae of the myxomycete Physarum polycephalum". Biology of the Cell. 63 (2): 239–248. doi:10.1016/0248-4900(88)90061-5. PMID 3060203 – via Science Direct.
- "Sclerotium". Wikipedia. September 26, 2019. Retrieved December 5, 2019.
- Dee, Jennifer (1960). "A Mating-type System in an Acellular Slime-mould". Nature. 185 (4715): 780–781. Bibcode:1960Natur.185..780D. doi:10.1038/185780a0.
- Wheals, Alan (1970). "A homothallic strain of the myxomycete Physarum Polycephalum". Genetics. 66: 623–633.
- Clark and Collins (1976). "Studies on the Mating Systems of Eleven Species of Myxomycetes". American Journal of Botany. 63 (6): 783–789. doi:10.1002/j.1537-2197.1976.tb11867.x. JSTOR 2442036.
- Burland, Timothy (1981). "Analysis of development and growth in a mutant of Physarum polycephalum with defective cytokinesis". Developmental Biology. 85 (1): 26–38. doi:10.1016/0012-1606(81)90233-5. PMID 7250516.
- Nakagaki, Toshiyuki; Yamada, Hiroyasu; Tóth, Ágota (2000). "Intelligence: Maze-solving by an amoeboid organism". Nature. 407 (6803): 470. Bibcode:2000Natur.407..470N. doi:10.1038/35035159. PMID 11028990.
- Nakagaki, Toshiyuki; Kobayashi, Ryo; Nishiura, Yasumasa; Ueda, Tetsuo (November 2004). "Obtaining multiple separate food sources: Behavioural intelligence in Physarum plasmodium". Proceedings of the Royal Society B. 271 (1554): 2305–2310. doi:10.1098/rspb.2004.2856. PMC 1691859. PMID 15539357.
- Tero, Atsushi; Takagi, Seiji; Saigusa, Tetsu; Ito, Kentaro; Bebber, Dan P.; Fricker, Mark D.; Yumiki, Kenji; Kobayashi, Ryo; Nakagaki, Toshiyuki (January 2010). "Rules for biologically inspired adaptive network design" (PDF). Science. 327 (5964): 439–442. Bibcode:2010Sci...327..439T. doi:10.1126/science.1177894. PMID 20093467.
- Moseman, Andrew (22 January 2010). "Brainless slime mold builds a replica Tokyo subway". Discover Magazine. Retrieved 22 June 2011.
- Adamatzky, Andrew; Jones, Jeff (2010). "Road planning with slime mould: If Physarum built motorways it would route M6/M74 through Newcastle". International Journal of Bifurcation and Chaos. 20 (10): 3065–3084. arXiv:0912.3967. Bibcode:2010IJBC...20.3065A. doi:10.1142/S0218127410027568.
- Adamatzky, Andrew; Alonso-Sanz, Ramon (July 2011). "Rebuilding Iberian motorways with slime mould". Biosystems. 5 (1): 89–100. doi:10.1016/j.biosystems.2011.03.007. PMID 21530610.
- Saigusa, Tetsu; Tero, Atsushi; Nakagaki, Toshiyuki; Kuramoto, Yoshiki (2008). "Amoebae Anticipate Periodic Events" (PDF). Physical Review Letters. 100 (1): 018101. Bibcode:2008PhRvL.100a8101S. doi:10.1103/PhysRevLett.100.018101. hdl:2115/33004. PMID 18232821.
- Barone, Jennifer (9 December 2008). "Top 100 stories of 2008 #71: Slime molds show surprising degree of intelligence". Discover Magazine. Retrieved 22 June 2011.
- Dussutour, Audrey; Latty, Tanya; Beekman, Madeleine; Simpson, Stephen J. (2010). "Amoeboid organism solves complex nutritional challenges". PNAS. 107 (10): 4607–4611. Bibcode:2010PNAS..107.4607D. doi:10.1073/pnas.0912198107. PMC 2842061. PMID 20142479.
- Bonner, John Tyler (2010). "Brainless behavior: A myxomycete chooses a balanced diet". PNAS. 107 (12): 5267–5268. Bibcode:2010PNAS..107.5267B. doi:10.1073/pnas.1000861107. PMC 2851763. PMID 20332217.
- Becchetti, Luca; Bonifaci, Vincenzo; Dirnberger, Michael; Karrenbauer, Andreas; Mehlhorn, Kurt (2013). Physarum Can Compute Shortest Paths: Convergence Proofs and Complexity Bounds (PDF). ICALP. Lecture Notes in Computer Science. 7966. pp. 472–483. doi:10.1007/978-3-642-39212-2_42. ISBN 978-3-642-39211-5.
- Caleffi, Marcello; Akyildiz, Ian F.; Paura, Luigi (2015). "On the Solution of the Steiner Tree NP-Hard Problem via Physarum BioNetwork" (PDF). IEEE/ACM Transactions on Networking. PP (99): 1092–1106. doi:10.1109/TNET.2014.2317911.
- Dirnberger, Michael; Neumann, Adrian; Kehl, Tim (2015). "NEFI: Network Extraction From Images". Scientific Reports. 5 (15669): 15669. arXiv:1502.05241. Bibcode:2015NatSR...515669D. doi:10.1038/srep15669. PMC 4629128. PMID 26521675.
- Adamatzky, Andrew (2010). Physarum Machines: Computers from Slime Mould. World Scientific Series on Nonlinear Science, Series A. 74. World Scientific. ISBN 978-981-4327-58-9. Retrieved 31 October 2010.
- Andrew, Adamatzky (2010). "Slime mould logical gates: Exploring ballistic approach". Applications, Tools, and Techniques on the Road to Exascale Computing. IOS Press. 2012: 41–56. arXiv:1005.2301. Bibcode:2010arXiv1005.2301A.
- Adamatzky, Andrew (6 August 2008). "Steering plasmodium with light: Dynamical programming of Physarum machine". arXiv:0908.0850 [nlin.PS].
- Adamatzky, Andrew (31 May 2011). "On attraction of slime mould Physarum polycephalum to plants with sedative properties". Nature Precedings. doi:10.1038/npre.2011.5985.1.
- Night, Will (17 May 2007). "Bio-sensor puts slime mould at its heart". New Scientist. Retrieved 22 June 2011.
- Night, Will (13 February 2006). "Robot moved by a slime mould's fears". New Scientist. Retrieved 22 June 2011.
Sources
- Gawlitta, W.; Wolf, K.V.; Hoffmann, H.U.; Stockem, W. (1980). "Studies on microplasmodia of Physarum polycephalum". Cell and Tissue Research. 209 (1): 71–86. doi:10.1007/bf00219924. PMID 7191783.
- Henry Stempen; Steven L. Stevenson (1994). Myxomycetes. A Handbook of Slime Molds. Timber Press. ISBN 978-0-88192-439-8.
External links
| Wikimedia Commons has media related to Physarum polycephalum. |
- Dussutour, Audrey. Talk given by a French slime mold specialist (video). TEDx – via youtube. (in French, with English subtitles available)
- "PhysarumPlus".
An internet resource for students of Physarum polycephalum and other a-cellular slime molds
- Ball, Philip (2008). "Cellular memory hints at the origins of intelligence". Nature. 451 (7177): 385. Bibcode:2008Natur.451..385B. doi:10.1038/451385a. PMID 18216817.
The learning and memory potential of Physarum polycephalum
- Time-lapse video of Physarum growing in a petri-dish (video) – via youtube.
- "Slime Mould Collective".
An international network of/for intelligent organisms
- "NEFI".
Software that can be used to extract networks from images of Physarum.