Hepcidin
Hepcidin is a protein that in humans is encoded by the HAMP gene. Hepcidin is a key regulator of the entry of iron into the circulation in mammals.[4]
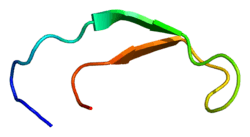
| Hepcidin | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Solution structure of hepcidin-25.[3] | |||||||||
| Identifiers | |||||||||
| Symbol | Hepcidin | ||||||||
| Pfam | PF06446 | ||||||||
| InterPro | IPR010500 | ||||||||
| SCOPe | 1m4f / SUPFAM | ||||||||
| OPM superfamily | 153 | ||||||||
| OPM protein | 1m4e | ||||||||
| |||||||||
| hepcidin antimicrobial peptide | |
|---|---|
| Identifiers | |
| Symbol | HAMP |
| NCBI gene | 57817 |
| HGNC | 15598 |
| OMIM | 606464 |
| RefSeq | NM_021175 |
| UniProt | P81172 |
| Other data | |
| Locus | Chr. 19 q13.1 |
During conditions in which the hepcidin level is abnormally high, such as inflammation, serum iron falls due to iron trapping within macrophages and liver cells and decreased gut iron absorption. This typically leads to anemia due to an inadequate amount of serum iron being available for developing red blood cells. When the hepcidin level is abnormally low such as in hemochromatosis, iron overload occurs due to increased ferroportin mediated iron efflux from storage and increased gut iron absorption.
Structure
Hepcidin exists as a preprohormone (84 amino acids), prohormone (60 amino acids), and hormone (25 amino acids). Twenty- and 22-amino acid metabolites of hepcidin also exist in the urine. Deletion of 5 N-terminal amino acids results in loss of function. The conversion of prohepcidin to hepcidin is mediated by the prohormone convertase furin.[5] This conversion may be regulated by alpha-1 antitrypsin.[6]
Hepcidin is a tightly folded polypeptide with 32% beta sheet character and a hairpin structure stabilized by 4 disulfide bonds. The structure of hepcidin has been determined through solution NMR.[3] NMR studies showed a new model for hepcidin: at ambient temperatures, the protein interconverts between two conformations, which could be individually resolved by temperature variation. The solution structure of hepcidin was determined at 325 K and 253 K in supercooled water. X-ray analysis of a co-crystal with Fab revealed a structure similar to the high-temperature NMR structure.[7]
Function
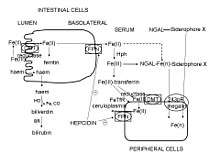
Hepcidin is a regulator of iron metabolism. Hepcidin inhibits iron transport by binding to the iron export channel ferroportin which is located on the basolateral surface of gut enterocytes and the plasma membrane of reticuloendothelial cells (macrophages). Hepcidin ultimately breaks down the transporter protein in the lysosome. Inhibiting ferroportin prevents iron from being exported and the iron is sequestered in the cells.[8][9] By inhibiting ferroportin, hepcidin prevents enterocytes from allowing iron into the hepatic portal system, thereby reducing dietary iron absorption. The iron release from macrophages is also reduced by ferroportin inhibition. Increased hepcidin activity is partially responsible for reduced iron availability seen in anemia of chronic inflammation, such as renal failure.[10]
Any one of several mutations in hepcidin result in juvenile hemochromatosis. The majority of juvenile hemochromatosis cases are due to mutations in hemojuvelin.[11] Mutations in TMPRSS6 can cause anemia through dysregulation of Hepcidin.[12]
Hepcidin has strong antimicrobial activity against E.coli ML35P N.cinerea and weaker antimicrobial activity against S.epidermidis, S.aureus and Group B streptococcus bacteria. Active against the fungus C.albicans. No activity against P.aeruginosa.[13]
Regulation
Hepcidin synthesis and secretion by the liver is controlled by iron stores within macrophages, inflammation, hypoxia, and erythropoiesis. Macrophages communicate with the hepatocyte to regulate hepcidin release into the circulation via eight different proteins: hemojuvelin, heriditrary hemochromatosis protein, transferrin receptor 2, bone morphogenic protein 6 (BMP6), matriptase-2, neogenin, BMP receptors, and transferrin.[14]
Erythroferrone, produced in erythroblasts, has been identified as inhibiting hepcidin and so providing more iron for hemoglobin synthesis in situations such as stress erythropoiesis.[15][16]
Vitamin D has been shown to decrease hepcidin, in cell models looking at transcription and when given in big doses to human volunteers. Optimal function of hepcidin may be predicated upon the adequate presence of vitamin D in the blood.[17]
History
The peptide was initially named LEAP-1, for Liver-Expressed Antimicrobial Protein, when it was first described in the year 2000.[18] Later, a peptide associated with inflammation was discovered, and named "hepcidin" after it was observed that it was produced in the liver ("hep-") and appeared to have bactericidal properties ("-cide" for "killing").[19] Although it is primarily synthesized in the liver, smaller amounts are synthesised in other tissues such as fat cells.[20]
Hepcidin was first discovered in human urine and serum in 2000.[21]
Soon after this discovery, researchers discovered that hepcidin production in mice increases in conditions of iron overload as well as in inflammation. Genetically modified mice engineered to overexpress hepcidin died shortly after birth with severe iron deficiency, again suggesting a central and not redundant role in iron regulation. The first evidence that linked hepcidin to the clinical condition known as the anemia of inflammation came from the lab of Nancy Andrews in Boston when researchers looked at tissue from two patients with liver tumors with a severe microcytic anemia that did not respond to iron supplements. The tumor tissue appeared to be overproducing hepcidin, and contained large quantities of hepcidin mRNA. Removing the tumors surgically cured the anemia.
Taken together, these discoveries suggested that hepcidin regulates the absorption of iron into the body.
Clinical significance
There are many diseases where failure to adequately absorb iron contributes to iron deficiency and iron deficiency anaemia. The treatment will depend on the hepcidin levels that are present, as oral treatment will be unlikely to be effective if hepcidin is blocking enteral absorption, in which cases parenteral iron treatment would be appropriate. Studies have found that measuring hepcidin would be of benefit to establish optimal treatment,[22] although as this is not widely available, C-reactive protein (CRP) is used as a surrogate marker.
β-thalassemia, one of the most common congenital anemias, arises from partial or complete lack of β-globin synthesis. Excessive iron absorption is one of the main features of β-thalassemia and can lead to severe morbidity and mortality. The serial analyses of β-thalassemic mice indicate hemoglobin levels decreases over time, while the concentration of iron in the liver, spleen, and kidneys markedly increases. The overload of iron is associated with low levels of hepcidin. Patients with β-thalassemia also have low hepcidin levels. The observations led researchers to hypothesize that more iron is absorbed in β-thalassemia than is required for erythropoiesis. Increasing expression of hepcidin in β-thalassemic mice limits iron overload, and also decreases formation of insoluble membrane-bound globins and reactive oxygen species, and improves anemia.[23] Mice with increased hepcidin expression also demonstrated an increase in the lifespan of their red cells, reversal of ineffective erythropoiesis and splenomegaly, and an increase in total hemoglobin levels. From these data, researchers suggested that therapeutics to increase hepcidin levels or act as hepcidin agonists could help treat the abnormal iron absorption in individuals with β-thalassemia and related disorders.[24] In later studies in mice,[25] erythroferrone has been suggested to be the factor that is responsible for the hepcidin suppression. Correcting hepcidin and iron levels in these mice did not improve their anemia.
References
- GRCh38: Ensembl release 89: ENSG00000105697 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- PDB: 1M4F; Hunter HN, Fulton DB, Ganz T, Vogel HJ (October 2002). "The solution structure of human hepcidin, a peptide hormone with antimicrobial activity that is involved in iron uptake and hereditary hemochromatosis". J. Biol. Chem. 277 (40): 37597–603. doi:10.1074/jbc.M205305200. PMID 12138110.
- Ganz T (August 2003). "Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation". Blood. 102 (3): 783–8. doi:10.1182/blood-2003-03-0672. PMID 12663437.
- Valore EV, Ganz T (2008). "Posttranslational processing of hepcidin in human hepatocytes is mediated by the prohormone convertase furin". Blood Cells Mol. Dis. 40 (1): 132–8. doi:10.1016/j.bcmd.2007.07.009. PMC 2211381. PMID 17905609.
- Pandur E, Nagy J, Poór VS, Sarnyai A, Huszár A, Miseta A, Sipos K (April 2009). "Alpha-1 antitrypsin binds preprohepcidin intracellularly and prohepcidin in the serum". FEBS J. 276 (7): 2012–21. doi:10.1111/j.1742-4658.2009.06937.x. PMID 19292870.
- PDB: 3H0T; Jordan JB, Poppe L, Haniu M, Arvedson T, Syed R, Li V, Kohno H, Kim H, Schnier PD, Harvey TS, Miranda LP, Cheetham J, Sasu BJ (September 2009). "Hepcidin revisited, disulfide connectivity, dynamics, and structure". J. Biol. Chem. 284 (36): 24155–67. doi:10.1074/jbc.M109.017764. PMC 2782009. PMID 19553669.
- Rossi E (August 2005). "Hepcidin--the iron regulatory hormone". Clin Biochem Rev. 26 (3): 47–9. PMC 1240030. PMID 16450011.
- Gulec S, Anderson GJ, Collins JF (August 2014). "Mechanistic and regulatory aspects of intestinal iron absorption". American Journal of Physiology. Gastrointestinal and Liver Physiology. 307 (4): G397–409. doi:10.1152/ajpgi.00348.2013. PMC 4137115. PMID 24994858.
- Ashby DR, Gale DP, Busbridge M, Murphy KG, Duncan ND, Cairns TD, Taube DH, Bloom SR, Tam FW, Chapman RS, Maxwell PH, Choi P (May 2009). "Plasma hepcidin levels are elevated but responsive to erythropoietin therapy in renal disease". Kidney Int. 75 (9): 976–81. doi:10.1038/ki.2009.21. PMID 19212416.
- Core AB, Canali S, Babitt JL (2014). "Hemojuvelin and bone morphogenetic protein (BMP) signaling in iron homeostasis". Frontiers in Pharmacology. 5: 104. doi:10.3389/fphar.2014.00104. PMC 4026703. PMID 24860505.
- Iron-Deficiency Anemia: New Insights for the Healthcare Professional: 2011 Edition. Scholarly Media LLC. Dec 2012. ISBN 978-1-4649-8960-5.
- "Hepcidin P81172". UniProt. December 15, 1998.
- Zhao N, Zhang AS, Enns CA (2013). "Iron regulation by hepcidin". J Clin Invest. 123 (6): 2337–43. doi:10.1172/JCI67225. PMC 3668831. PMID 23722909.
- Koury, M.J. "Erythroferrone: A Missing Link in Iron Regulation". The Hematologist. American Society of Hematology. Retrieved 26 August 2015.
- Kautz L, Jung G, Valore EV, Rivella S, Nemeth E, Ganz T (Jul 2014). "Identification of erythroferrone as an erythroid regulator of iron metabolism". Nature Genetics. 46 (7): 678–84. doi:10.1038/ng.2996. PMC 4104984. PMID 24880340.
- Bacchetta J, Zaritsky JJ, Sea JL, Chun RF, Lisse TS, Zavala K, Nayak A, Wesseling-Perry K, Westerman M, Hollis BW, Salusky IB, Hewison M (2014). "Suppression of iron-regulatory hepcidin by vitamin D". J. Am. Soc. Nephrol. 25 (3): 564–72. doi:10.1681/ASN.2013040355. PMC 3935584. PMID 24204002.
- Krause A, Neitz S, Mägert HJ, Schulz A, Forssmann WG, Schulz-Knappe P, Adermann K (September 2000). "LEAP-1, a novel highly disulfide-bonded human peptide, exhibits antimicrobial activity". FEBS Lett. 480 (2–3): 147–50. doi:10.1016/S0014-5793(00)01920-7. PMID 11034317.
- Park CH, Valore EV, Waring AJ, Ganz T (March 2001). "Hepcidin, a urinary antimicrobial peptide synthesized in the liver". J. Biol. Chem. 276 (11): 7806–10. doi:10.1074/jbc.M008922200. PMID 11113131.
- Bekri S, Gual P, Anty R, Luciani N, Dahman M, Ramesh B, Iannelli A, Staccini-Myx A, Casanova D, Ben Amor I, Saint-Paul MC, Huet PM, Sadoul JL, Gugenheim J, Srai SK, Tran A, Le Marchand-Brustel Y (September 2006). "Increased adipose tissue expression of hepcidin in severe obesity is independent from diabetes and NASH". Gastroenterology. 131 (3): 788–96. doi:10.1053/j.gastro.2006.07.007. PMID 16952548.
- Kemna EH, Tjalsma H, Willems HL, Swinkels DW (January 2008). "Hepcidin: from discovery to differential diagnosis". Haematologica. 93 (1): 90–7. doi:10.3324/haematol.11705. PMID 18166790.
- Bregman DB, Morris D, Koch TA, He A, Goodnough LT (February 2013). "Hepcidin levels predict nonresponsiveness to oral iron therapy in patients with iron deficiency anemia". Am. J. Hematol. 88 (2): 97–101. doi:10.1002/ajh.23354. PMID 23335357.
- Gardenghi S, Ramos P, Marongiu MF, Melchiori L, Breda L, Guy E, Muirhead K, Rao N, Roy CN, Andrews NC, Nemeth E, Follenzi A, An X, Mohandas N, Ginzburg Y, Rachmilewitz EA, Giardina PJ, Grady RW, Rivella S (December 2010). "Hepcidin as a therapeutic tool to limit iron overload and improve anemia in β-thalassemic mice". J. Clin. Invest. 120 (12): 4466–77. doi:10.1172/JCI41717. PMC 2993583. PMID 21099112.
- Kroot JJ, Tjalsma H, Fleming RE, Swinkels DW (December 2011). "Hepcidin in human iron disorders: diagnostic implications". Clin. Chem. 57 (12): 1650–69. doi:10.1373/clinchem.2009.140053. PMID 21989113.
- Moura IC, Hermine O (2015). "Erythroferrone: the missing link in β-thalassemia?". Blood. 126 (17): 1974–5. doi:10.1182/blood-2015-09-665596. PMID 26494918.
Further reading
- Camaschella C (2005). "Understanding iron homeostasis through genetic analysis of hemochromatosis and related disorders". Blood. 106 (12): 3710–7. doi:10.1182/blood-2005-05-1857. PMID 16030190.
External links
- hepcidin at the US National Library of Medicine Medical Subject Headings (MeSH)
- Intrinsic LifeSciences - Hepcidin Research Facility, The BioIron Company
- Hepcidinanalysis.com - Service for Hepcidin measurements: Scientific Research, Patients and Clinical Trials
- Protein Data Bank Page
- PDBe-KB provides an overview of all the structure information available in the PDB for Human Hepcidin